![]()
ON THE NATURAL EQUILIBRIUM OF PYRAUSTA NUBILALIS HB.(Suite)
![]()
The
immunity reaction of the host larvae against the eggs of Limnerium alkae
destroys a very large percentage of the parasites. Another very important factor,
resulting in a high mortality, is the usual deposit of several eggs in the same
host larva. It is well known that only one parasite larva develops in the same
host larva. The hatching of the first parasite means death to the rest. Their
death does not, however, as THOMPSON & PARKER believe, result from the
action of a cytolytic enzyme appearing in the blood of the host after the first
parasite larva has hatched.
The
importance of the mortality of the parasites in the egg stage is, illustrated in
Table 1. The observations were made in the field on Corn Borer larvae in various
stages of development. It may be concluded from the data, presented in the
table, that the Limnerium alkae larvae,. which successfully complete their
development, only represent a small proportion of the deposited eggs. The
mortality varies considerably from one corn field to another. In the 8 series of
observations listed in the. tables, the mortality was 91, 83, 82, 71, 84, 88,
92, and 96 per cent. respectively. The Corn Borer does not seem to be a host,
favorable to the propagation of Limnerium alkae.
When
the larva of Limneriuni alkae hatches, it possesses a tail and its head is
protected by a chitinous hood (Fig. 7). Its form is identical with that of
Lininerhim crassifemur. The tail and the hood is lost at the first moult.
Anatomically,
there are slight differences between the larvae of the two species. The
malphigian tubes are decidedly longer in Limnerium alkae than in Lininerium
crassifemur. In the former species they are longer than one-third of the length
of the body (without the tail), in the latter species they are less than one
fifth of the length of the body.
The
difference is still more pronounced in the older larvae. The malphigian
apparatus is composed of two straight double vessels, one on each side of the
alimentary canal. One branch of each double vessel is slightly longer than the
other. The vessels open through a very short common tribe into a rectal
reservoir without direct connection with the intestines.
Fig. 7, 8 et 9
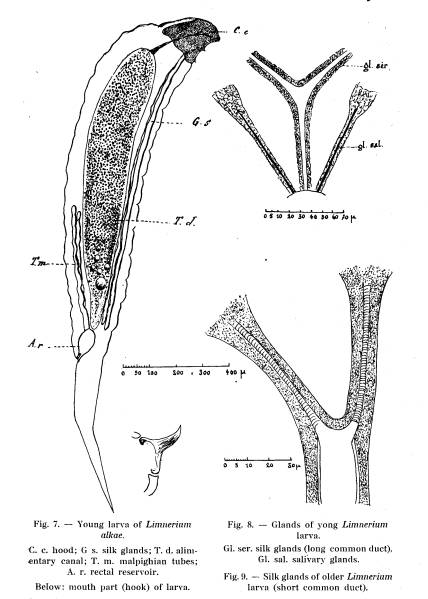
The two ducts of the salivary glands and the common trunk of the silk
glands open at the mouth. The salivary glands are composed of two relatively
short tubes, one on each side of the head. They consist of large, granular cells.
The silk glands form two double tubes along the sides of the alimentary canal.
The two tubes fuse to a common duct opening ventrally in the mouth. The common
ductis decidedly longer in the primary larvae (Fig. 8) than in those which have
lost tail and hood. In the latter, however, the duct is much wider than in the
very young larvae (Fig. 9). The walls of the silk gland tubes are transversally
striated due to a spiral thickening of the chitin very similar to that of the
tracheae.
On
each side of the dorsal blood vessel, groups of granular pericardial cells are
located. When the larvae are placed in physiological salt solution to which is
added neutral red, the granules absorb the color very readily. The nervous
system is composed of oblong ganglia; the longitudinal cords connect directly
with the brain.
The
mouth parts consist of two chitinous hooks, very similar to those of the larva
of Limnerium crassifemur (Fig. 7). The very pointed hooks are curved, and move
from side to side between two lateral .chitinous pieces, a relatively long one
behind and a short one in front. The connection with the hind piece is similar
to that described for Limnerium crassifemur. The first larva hatched in each
host kill those hatched later with these pointed hooks.
Limnerium
alkae hibernates in the same way as Lininerium crassifemur. The full grown larva
spins a very resistant grayish cocoon beside the remnants of the dead host. The
pupa resembles that of Limnerium crassifemur.
In
conclusion it may be stated, that Lininerium alkae, although found in the
majority of the infested corn fields, is poorly adapted to the Corn Borer. Its
spread seems to depend upon the presence of an intermediary host which, in fact,
might be considered the principal host. The existence of that host is still
hypothetical. Future researches ought to elucidate this obscure point in the
biology of the parasite.
MICROBRACON
BREVICORNIS Wesmael.
The
spread of this parasite is very irregular in the investigated territory. It
attacks the full grown Corn Borers, paralyzes them with a sting and deposits the
eggs, usually in masses, on the skin of the host. GENIEYS has made a very
complete study of the biology of this species. I have been able to verify the
majority of his observations. I have, however, noted that the number of eggs
deposited on each host rarely exceeds 10.
The
female of Alicrobracon does not always oviposit on the paralyzed larvae.
Paralyzed but not parasitized Corn Borers are frequently found in the corn
stems. They strongly resemble the larvae killed by bacteria.
The
development of Microbracon is very rapid but varies with the temperature. In the
year 1927, when the summer temperature was below normal, the development of the
parasite was decidedly slower than in 1928. On September 1, 1927, a parasitized
Corn Borer, carrying 9 Microbracon eggs, was brought to the laboratory. The
parasite larvae hatched on September 2, commenced spinning their cocoons on
September 7, and pupated September 10. The adults emerged on September 21. In
1928, when the mid-summer was exceptionally warm, a Microbracon female deposited
two eggs on a Corn Borer on August 23. The larvae appeared during the night
August 24-25. On August 27 thelarvae had reached their maximum size and stopped
eating. Within 14 hours they left the host and began spinning their cocoons. The
adults emerged on September 6. The metamorphosis lasted but 14 days compared
with 22 days the previous year, at approximately the same time.
The
rapid development, and the fact that Microbracon is better adapted to the Corn
Borer than is Limnerium alkae, favors the propagation of the parasite.
Nevertheless it does not multiply more rapidly than Limnerium alkae. Several
factors limit its spread. THOMPSON & PARKER mention the following: its
polyphagous habits, its low fertility, the difficulty of entering into the
interior of the corn stalks, and the fact that each female only parasitizes one
or a few Corn Borers.
In
certain corn fields of the Jura region, the mortality of the Corn Borers due to
Microbracon brevicornis amounted to 10 per cent. during 1927. This is
exceptional however, the average being barely 5 per cent.
LYDELLA
SENILIS Meigen.
The
distribution of this fly is more limited than the distribution of Limnerium
alkae. I have found it in the Rhone valley, especially at Lyon, and a few
specimens have been collected in material from the Jura. It does not seem,
however, that the parasite plays an appreciable rôle in the latter region.
The
following observations indicate the importance of Lydella senilis in the region
where it is most plentiful. In 1927, 106 Corn Borers were examined in a corn
field 30 kin north-east of Lyon. Of these, 5 were parasitized by Lydella, one by
Limizerium. In another field near by, 105 Corn Borers were examined, of which 15
were parasitized by Lydella and none by Limnerium. Two other surveys in this
region have shown that the degree of parasitism by Lydella fluctuates between 5
and 10 per cent. In this particular region, it is always higher than the
parasitism by Limnerium.
Ordinarily
only one parasite larva is found in each host. In 1928, however, I have twice
found a Corn Borer containing two Lydella larvae. The two parasites in the same
host do not seem to disturb one another, but they are smaller than solitary
larvae. The adults, also, become smaller than the normal type. It is possible
that Ludella senilis has some other host besides the Corn Borer. The following
observation seems to substantiate this. In the beginning of the month of July,
1927, I placed some Corn Borer eggs, collected in south-western France, on corn
plants grown in the garden of the Entomological Station. At the end of July, I
found five Lydella pupae besides dead Corn Borer larvae. The parasites probably
came from some other host, since the station is located far from any corn fields
infested by the Corn Borer, .and no corn had been grown in the garden for
several years. The :spread of Lydella senilis probably depends upon the presence
of intermediary hosts. Unfortunately these are not known.
OCCASIONAL
CORN BORER PARASITES
II
-- BACTERIA
Several
bacteria have been isolated in 1927 from dead Corn Borers and from those
paralyzed by Microbracon brevicornis. Tree coccobacillae and one Gram positive
micrococcus were obtained in pure culture and used in laboratory and field
experiments to infect Corn Borers per os.
In
the laboratory experiments, corn stalk pieces were submerged in an emulsion of
bacteria in physiological salt solution and afterwards kept in petri-glasses.
Corn Borer larvae of different sizes were placed in the glasses. They soon bored
their way into the stalk pieces, thereby absorbing a considerable number of
bacteria. The experiments were made on August 12, 1927 with coccobacillae and
the micrococcus, isolated on August 5 and 6 in the Jura. No positive results
were obtained.
Similar
experiments were made on the same day with two sporeforming bacteria, pathogenic
for the silk worm; Ischivata's Bacillus sotto, by the Japanese authors
erroneously considered the cause of
the flacherie, and a bacterium sent by Professor M. G. BALERIOLA Of Valencia,
Spain. The latter species is so virulent, that it kills the silk worms in less
than 24 hours. Three days later, three of the ten Corn Borers infected by
Bacillus sotto, showed signs of disease, while the others were still healthy.
The Spanish bacterium had no effect on the Corn Borers.
In
the field experiments, some drops of the emulsion were placed at the base of the
corn plant leaves. They contaminated the region between the leaf and the stem
where Corn Borers, placed on the plant, usually bore through the stem. The
experiments were made at Toulouse (Jura) on August 13, 1927. Three rows of six
corn plants each were infected with two coccobacillae and a micrococcus
respectively. No positive results were obtained.
I
have not yet been able to find diseases caused by filterable viruses. Such
diseases, because of their hereditary character, would be more useful than
bacterial diseases.
III
- PROTOZOA
Protozoa
play a much more important part than bacteria in the natural destruction of
noxious insects. Most of the forms, pathogenic to insects, belong to the group,
Microsporidia. Flagellates are also frequent in insects, but their parasitism is
less pronounced. Many of them live in the insects as commensals. The
Microsporidia, like the filterable viruses, live in the interior of the cells
and may be transmitted from one generation to the other through the egg. They
are not as dependent upon favorable outside factors as are bacteria and
especially fungi. For that reason, they are the most valuable auxilliary
microorganisms, from an agricultural standpoint.
PEREZIA PYRAUSTAE nov. sp.
So
far, I have only found one protozoon of the group, Microsporidia, in the Corn
Borer. It is closely related to the one I discovered in 1919 in the malpighian
vessels of the larvae of Pieris brassicae and named Perezia mesnili. The Corn
Borer parasite, Perezia pyraustae, lives in cells of the malpighian vessels and
of the silk glands of the host larvae. The parasitized larvae cannot be
distinguished from the normal ones by any outward symptom, not even by means of
a magnifying glass.
When
a parasitized larvae is dissected and the organs examined with binocular
microscope, the abnormal development of the malpighian vessels is striking (Fig.
10). These vessels are normally fine tubes, transparent towards the distal end
and increasingly opaque and yellow towards the opening into the alimentary
canal. When infected, the vessels become opaque throughout their length and turn
more and more white. At the same time they swell. The yellow colour of their
basal part is retained.
By
examining a fresh fragment of the white and -hypertrophied part of the tube
under the microscope, the majority of the cells are seen to be filled with
ovoform refractive bodies (Fig. 11). These bodies are the spores of the
parasite, they are smaller than the spores of the related microorganism causing
pebrine among silk worms. The parasitized cells of the vessel walls are
hypertrophid. The ciliated epithelium towards the lumen of the vessels is
destroyed where the parasites are numerous.
A
study of the live cycle of Perezia pyraustae was made on film preparations
stained with Giemsa. The pictures, obtained by this simple and rapid method, are
exactly identical with those observed with considerable difficulty on fresh
material or on microtome sections.
The
asexual phase of the development of the parasite is represented by small round
cells, the protoplasma of which stain deep blue with Giesma (Fig. 12). They
usually contain two nuclei, but sometimes peculiar nuclear elements, looking
like clearly separated chromatine grantiles, may be observed. The shape of the
nuclei is rather variable. Each of the nuclear types, illustrated in Fig. 12,
probably represents a particular phase in the development of the parasite, but
it, is impossible to determine their exact relations and individual significance..
The division of the parasite takes place in one or several directions.. In the
first case small chains. of binuclear cells result; in the other case
multinuclear elements are formed. The small chains rarely con tain more than two
cells, and the multinuclear elements never show more than four nuclei. In
related forms, the number of nuclei often reach eight.
When
the living conditions of the cells become less favorable, and especially when
their cytoplasm becomes entirely filled up with parasites, the development is
considerably modified (Fig. 13). The parasites become more oblong and their
protoplasm develops vacuoles and stains pale blue with Giemsa. The nuclear
structure changes at the same time. Some parasites show a more or less- regular
group of chromatin granules (Fig. 13 nos. 2 and 3); others form a chromatine
mass of indefinite form (no. 1); others show two separate groups of granules
(no. 9); some have four nuclei arranged in pairs (nos. 6-8). The cells with four
nuclei are the sporoblasts which, form the spores through division. The
processes, which result in the formation of spores, can not be observed on the
stained film preparations. Only two of the nuclei stain well. They occupy a
central position and consist of condensed chromatin. I have not, by any of the
appropriate means, been able to observe the formation of polar bodies.
The
size of the spores differs somewhat in the different hosts. Sometimes double
spores and oversized spores occur. Certain hibernating Corn Borers, sent to me
from the departments of Aude and Tarn contained a particularly large number of
double spores and oversized spores.
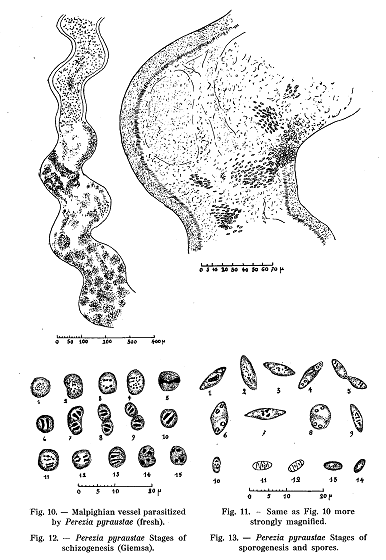
The
sporoblasts. giving rise to two spores, are characteristic for the Corn Borer
parasite of the group Microsporidia. For that reason it is. natural to refer it
to the genus Perezia to which belong three species found in the larvae of Pieris
brassicae. The existence of double spores is another argument in favor of this
placing. The normal spores of the Corn Borer parasite are very similar to those
of Perezia mesnili. I believe, however, that it is proper to consider them as
two different species and to describe the Corn Borer parasite as a new species
Perezia pyraustae.
It is
very easy to infect Corn Borer larvae with the parasite. It suffices to crush
the malphigian vessels of an infected larva in sterile physiological salt
solution, to moisten corn stalk pieces with the fluid and give them as feed for
healthy Corn Borers. When these bore their way into the stalk pieces they absorb
enough spores to start a new infection. The spores germinate, and the ameboid
parasite moves into the malphigian vessels, or the spores pass directly into the
vessels or in the
Perezia pyraustae has
been found in the Jura, and especially in the regions of Bletterans and Chaussin
which seem to be two important centers for the spread of the disease. The
percentage of infestation, in 1927, reached 30 and in some fields 40. Around
these centres, the degree of infestation is less. In the corn fields south of a
line Lons-le-Saulnier - Louhans, I have never found a single
infected larva, although I have examined thousands. The parasite has been found
in the Corn Borer material from the departments of Tarn and Aude, but I have no
exact data on its distribution in those territories.
Perezia pyraustae, although
rather abundant in France, does not seem to be of much consequence in the
natural destruction of the Corn Borer. It is possible, however, that the disease,
which attacks the Corn Borer in all developmental stages, reduces its vitality
and favors the action of other destructive factors.
![]()
![]()