Nos chromosomes sont constitués
de deux parties bien différentes.
La plus petite comprend des séquences
qui codent les protéines et que l'on appelle les gènes. À
lire les journaux ou à écouter les radios et télévisions,
nul ne peut ignorer que "tout est dans les gènes".
L'autre
partie, c'est-à-dire les 3/4 au moins de leur longueur, ne porte
aucun code. Pour cette raison, certains appellent ces séquences
du "chromosome poubelle".
À l'intérieur de cette
croyance très répandue selon laquelle toute la construction
d'un être vivant est définie par le code porté par
ses gènes, il restait cependant une zone d'ombre : toutes les cellules
de notre corps comportent les mêmes gènes, pourtant, elles
sont bien différentes les unes des autres et ne font pas la même
chose.
Comment cela est-il possible ?
On se disait que pour ce faire,
le code génétique contient nécessairement des instructions
qui disent comment l'utiliser différemment dans des circonstances
différentes, et l'on supposait qu'au fur et à mesure de la
croissance d'un organisme chaque type de cellule spécialise son
code d'une façon ou d'une autre.
On se le disait, jusqu'à
ce qu'une certaine brebis Dolly soit clonée à partir d'une
cellule supposée s'être ainsi spécialisée pour
ne plus savoir faire que son métier de cellule de glande mammaire.
Or,
sans que les généticiens soient le moins du monde intervenus
sur le code génétique de cette cellule, elle s'est montrée
capable tout à trac de générer un organisme entier
: ses gènes n'étaient donc pas spécialisés.
Pour parfaire la mise à mal
de la théorie du "Big-Gène qui fait tout", des experts du
CNRS et de l'Institut Curie viennent de trouver que les humains auraient
presque les mêmes gènes qu'un petit singe d'Amérique
du Sud qu'on appelle Capucin [revue SCIENCES ET AVENIR de juillet
1997] : même
la différence entre les humains et les singes ne serait donc pas
dans les gènes ! Où va-t-on ?
On va tout simplement à devoir
admettre la réalité : tout n'est pas dans les gènes.
L'essentiel même ne serait
pas dans les gènes, puisque les gènes de la cellule de glande
mammaire n'ont pas été modifiés entre le moment où
ils fonctionnaient en cellule mammaire et le moment où ils fonctionnaient
en cellule souche d'organisme entier, et puisque les gènes du capucin
ne le transforment pas en humain et inversement.
Si l'essentiel
n'est pas dans le code génétique, où est-il donc ?
Nous n'avons pas le choix :
l'essentiel
est nécessairement dans le chromosome "poubelle"
qui ne porte aucun code de protéine.
L'apparition de la vie : un scénario à l'envers
Usuellement, on suppose que des
cellules vivantes se sont crées de quelque façon mystérieuse,
puis que, pour transmettre efficacement les instructions utiles à
leur reproduction et à leur fonctionnement, les cellules ont inventé
le non moins mystérieux procédé de codage génétique
porté par la double hélice d'ADN que l'on appelle chromosome.
Comment ce code fonctionne-t-il
? Nul n'en s'est rien, mais on constate que la présence de telle
ou telle séquence génère la production de telle ou
telle protéine bien spécifique.
Notre hypothèse de départ
prend les choses à rebours : on
suppose que ce sont les chromosomes en ADN qui se sont formés les
premiers, puis, qu'ils ont tout simplement organisé une cellule autour
d'eux pour mieux fonctionner.
Quant au code génétique,
comme on le verra : on suppose qu'il ne fonctionne pas comme un code !
Les mouvements de la vie
= de simples mouvements électro-magnétiques
Cette hypothèse s'appuie
sur une idée simple : il n'y aurait aucune
différence entre les mouvements que l'on dit ceux de la vie et
les mouvements d'attirance et de répulsion sous l'effet des forces
magnétiques qui font bouger "toute seule" la matière inanimée.
Une cellule vivante ne serait rien
d'autre qu'un agencement de matière, tel que les mouvements d'origine
magnétique engendrés par ses atomes parviennent tout simplement
à se refermer en boucle autonome et incessante.
La
forme en hélice des chromosomes serait liée à leur
faculté de conduire un courant électrique et de se transformer
ainsi en aimant.
Le mouvement
de division des cellules s'expliquerait pour l'essentiel par la répulsion
magnétique d'aimants/chromosomes de même signe et imbriqués
l'un dans l'autre.
Dans cette conception, la forme
en double hélice des chromosomes est essentielle : un
chromosome ne serait fondamentalement qu'un circuit électrique enroulé
en hélice, et comme tout circuit électrique en forme d'hélice
il se comporte en aimant muni d'un pôle Nord et d'un pôle Sud.
La division cellulaire ne serait
alors que l'effet d'un gonflement suffisant du courant électrique
parcouru par le chromosome, de telle sorte que chacune des deux hélices
se comporte soudainement comme un aimant autonome. Dans cette situation
où deux aimants de même sens se retrouvent imbriqués,
très naturellement ils se repoussent mutuellement, déchirent
leurs liaisons, s'écartent tant qu'ils le peuvent, et créent
deux cellules séparées en reconstituant leurs doubles. Ces
doubles subiront le même sort lorsque, à nouveau, le courant
repassera avec suffisamment d'abondance.
additif d'octobre 2001 :
A la Colombia University de New-York, des recherches sont menées
concernant l'effet des champs électromagnétiques sur l'ADN
des cellules. Ces recherches montrent que de faibles champs parviennent
à initier la transcription de certaines protéines (notamment
de stress), en développant à des sites spécifiques
des forces de répulsion entre les deux brins d'ADN. Les chercheurs
pensent ces forces en terme de répulsion d'origine électrique,
liée à l'accélération par le champ électromagnétique
des électrons conduits par chaque brin de la double hélice.
Cette force de répulsion entre les deux brins occasionne leur séparation,
ce qui amorce alors le fonctionnement de l'ARN polymérase et la
transcription des protéines.
Ce mécanisme de répulsion d'origine électromagnétique
entre les deux brins de la double hélice concerne ici seulement
des séparations ponctuelles, mais il se rapproche beaucoup du mécanisme
de répulsion que l'on suppose à l'origine non seulement des
transcriptions locales de l'ADN, mais aussi de son mécanisme de
transcription complète lors de la division cellulaire.
Ces études sont principalement menées par Martin BLANK
et Reba GOODMAN, et on peut librement trouver sur le site de Wiley InterSience le
résumé de leur dernier article publié en mars 2001
(
accès à : Electromagnetic initiation of transcription at specific DNA sites). Depuis
la page de ce résumé, on peut obtenir d'autres résumés
de textes publiés par les mêmes auteurs.
Sur ce principe, on peut très
facilement expliquer que des molécules organisées en double
hélice se mettent à proliférer, se répliquant
sans fin à l'identique. Cette prolifération d'hélices
"vivantes" ne fait pas intervenir la moindre étincelle de vie en
supplément des règles connues des forces magnétiques
qui s'appliquent à la matière dite "morte".
Que ces chromosomes se groupent
ensuite en paquet, puis qu'ils organisent autour d'eux le réseau
fermé d'une cellule, n'est plus dans cette hypothèse qu'une
manière pour les chromosomes de s'organiser à plusieurs pour
que le courant électrique qui les fait vivre ne se disperse pas
en quittant l'extrémité de leur hélice, pour qu'il
puisse se recycler au maximum en interne : il sort par le bout d'un chromosome,
parcourt captif l'un des tubules qui forment le squelette du noyau, puis
repénètre dans un autre chromosome.
Construction
du réseau des tubules d'une cellule pour recycler en permanence
le courant électromagnétique parcourant un groupe de chromosomes
Signalons que la capacité
d'un chromosome à se comporter comme conducteur de courant a été
démontré de plusieurs manières :
- d'une part
il est devenu usuel d'observer l'ADN à l'aide de microscopes à
effet tunnel, or un tel microscope ne peut fonctionner que si la surface
observée est conductrice ou semi-conductrice.
- d'autre part,
la chimiste Jackie Barton de l'Institut californien de technologie à
Pasadena a mis en évidence le transport possible d'électrons
le long de la molécule d'ADN [voir SCIENCES ET AVENIR de juillet
1997]. Dans l'additif d'octobre 2001 ci-dessus, il est fait référence
à des recherches qui considèrent que ce transport d'électrons
par l'ADN est un fait maintenant acquis, sur la base d'expériences
menées par l'équipe de WAN en 1999 et de PORATH en 2000.
Les chromosomes ont une forme dans l'espace
À partir de cette idée
que les chromosomes ne seraient que des bobines électriques fonctionnant
comme des aimants, on se demande maintenant comment les cellules se spécialisent
selon les organes tout en portant le même code génétique.
Bien entendu, cela pose la question
:
Que vient donc faire le code génétique dans cette histoire ?
Le courant électromagnétique
qui circule dans les chromosomes est à prendre au sérieux
en tant que "courant", c'est- à-dire qu'il
se déroule avec une certaine forme dans l'espace, de la même
façon que le courant d'une rivière suit des méandres
géométriques précisément définis dans
une vallée.
Si une
protéine quelconque vient se prendre dans le filet du paquet de
chromosomes, sa présence déforme leur courant électrique,
elle l'oblige à se détourner d'une façon particulière
qui dépend de la forme de la protéine elle-même et
du courant magnétique qu'elle fait régner dans son entourage.
La
présence d'une protéine déforme le courant électrique
qui parcourt l'hélice des chromosomes et marque ainsi son empreinte
sur eux
L'ADN est constitué d'une
succession de deux paires de bases : A ou T, et G ou C.
On suppose que la paire complémentaire
A et T implique une certaine courbure au courant électrique qui
suit l'hélice d'ADN, tandis que la paire G et C lui implique une
autre courbure.
Pour connaître une courbure
régulière, il suffit d'en connaître trois points, puisque
par trois points on ne peut faire passer qu'une courbe régulière et une seule. Pour
cette raison, la forme exacte de la courbure du courant électrique
qui parcourt l'ADN peut être connue si on la décompose en
unités élémentaires suffisamment fines et correspondant
chacune à trois bases. Cette
succession de chapelets de trois, nous pouvons l'appeler "code génétique",
mais dans notre optique cela n'implique pas une succession d'instructions
en langage codé, mais une succession d'intensités de torsions
capable de décrire et de recréer la courbure du courant électrique
qui parcourt le chromosome.
Par
3 points on ne peut faire passer qu'une seule courbe.
Pour cette
raison, la courbure du courant des chromosomes peut se décomposer
en "triplets d'intensités de courbure".
Chaque
valeur d'intensité correspond à l'une des deux paires de
base du code génétique.
Le principe de la réplication des protéines :
Quand une protéine vient donc
se prendre dans le paquet des chromosomes, elle déforme son courant,
puis, quand le chromosome va se scinder en deux pour se recopier, ce sont
les paires de base correspondant au courant déformé qui vont
s'installer, et c'est ainsi un courant déformé par la présence
de la protéine qui va désormais se dupliquer.
Cette protéine laissera donc
comme une empreinte en creux dans le chromosome dupliqué, et le
chromosome ne retrouvera son équilibre électromagnétique
que lorsque ce creux en lui sera rempli par des matériaux aspirés
magnétiquement par ce défaut dans son courant. En le comblant,
ces morceaux reconstruiront involontairement une réplique exacte
de la protéine initiale.
Si l'organe où se trouve
la cellule a besoin de la protéine, il n'aura qu'à se servir,
et le trou à nouveau vacant servira de moule à une nouvelle
protéine.
On voit que dans ce fonctionnement,
le "code"
qui correspond à la séquence des bases le long de l'ADN ne
fonctionne pas comme un code, mais c'est bien lui qui implique la production
de telle ou telle protéine
plutôt qu'une autre.
On en revient donc à la question initiale :
Comment
ce système de recopie automatique d'une protéine se spécialise-t-il selon les organes
?
On suppose que cela se fait de la
manière la plus simple : pour
qu'une protéine soit spécialement utilisée et fabriquée
par une cellule, il suffit que son site se trouve bien exposé à
la périphérie du paquet des chromosomes.
Si au contraire la cellule n'a pas pour fonction de produire tel type de
protéine, son site sera enfoui au profond du paquet des chromosomes,
et il sera recouvert par des sites plus utiles à l'organe concerné.
Et comment une cellule fait-elle
donc pour enfouir spécialement une partie de ses sites de protéines,
et pour exposer spécialement en périphérie d'autres
sites qui lui sont plus utiles ?
La réponse semble aisée
: les
chromosomes se tordent, se faufilent les uns dans les autres, ou s'extraient
du paquet pour faire venir, avec eux, les sites concernés à sa périphérie.
Et qu'est-ce
qui donne sa forme à ce paquet
sans déformer les sites de protéines eux- mêmes ?
C'est
tout simplement le fameux "chromosome-poubelle",
qui lui aussi adapte ses bases pour que leur courbure change la conformation
dans l'espace du paquet des chromosomes, et pour qu'elle spécialise
ainsi dans chaque cellule les sites de protéines accessibles.
Cette
hypothèse s'accorde avec l'expérience Dolly,
car si les biologistes n'ont pas modifié le code génétique
de la cellule de glande mammaire, ils lui ont cependant fait subir un traitement
significatif : ils lui ont imprimé des décharges électriques.
Selon nous, ces décharges
ont eu pour effet d'emporter le paquet des chromosomes dans un fort courant
électrique, ce qui a défait les torsions locales du chromosome-poubelle
qui avaient spécialisé la cellule. Elles l'ont ainsi complètement
dé-spécialisée sans pour autant modifier son code
génétique.
Si l'observation sur l'identité
presque complète du génome humain et du génome capucin est
juste, alors ce ne serait pas seulement le type de protéine produit
qui est déterminé par la forme dans l'espace que l'agencement
du chromosome-poubelle donne à l'ensemble du paquet des chromosomes,
mais c'est toute l'évolution de la spécialisation des organes
qui serait induite de la même façon.
Dans un complément de 2008, disponible en ligne, il est relaté le résultat de recherches réalisées postérieurement à ce texte et qui confirment, à la fois l'importance du chromosome poubelle dans le rôle de spécialisation des cellules, et l'importance de l'accessibilité plus ou moins grande des sites de protéine comme moyen de cette spécialisation.
On n'échappe
pas aux mathématiques
Le principe du fonctionnement des
forces dans l'univers a pu être énoncé par Newton lorsqu'a
été maîtrisée la question mathématique
des dérivées. On pourrait croire que la question du vivant
et de son fonctionnement ne nécessiterait pas une réflexion
de type mathématique pour être pensée.
Il n'en est rien, car l'hypothèse
que l'on vient de faire et qui pourrait expliquer de façon simple
le fonctionnement des cellules vivantes, ne tient pas si l'on ne fait pas
une subtile modification dans nos conceptions mathématiques.
[la section F
"Mathématique" de ce site est intégralement
consacrée à l'exposition détaillée de cette
modification qu'il convient de faire dans nos conceptions]
C'est
qu'en effet l'organisation dans l'espace du chromosome, telle qu'on l'a
supposée pour expliquer son fonctionnement, ne s'observe pas dans
la réalité.
Dans la réalité, sauf
au moment de la division cellulaire où le chromosome se condense
sous la forme du bâtonnet que l'on représente habituellement,
le chromosome n'apparaît dans la cellule que comme un filament déroulé
et flageolant, et ne semble présenter aucune structure particulière
stable dans l'espace.
Le problème
mathématique revient à comprendre pourquoi et comment le
chromosome déroulé et flageolant que l'on voit dans la "vie
courante" de la cellule, fonctionne exactement comme s'il était
condensé en une forme spatiale stable.
C'est une question mathématique,
parce qu'elle concerne l'idée que l'on se fait des "dimensions"
de l'espace, et parce qu'elle met notamment en cause le nombre de ces dimensions.
Un phénomène tel qu'un
courant électrique peut-être soumis à plusieurs tendances
simultanées, c'est-à-dire à plusieurs forces distinctes
qui chacune tend à déformer ce courant de façon particulière.
Par exemple, l'une de ces forces peut tendre à l'étirer dans
un sens, une autre peut tendre à l'étaler vers une autre
direction, et une autre encore à le faire dévier systématiquement
d'un autre côté.
Comme l'espace possède trois
dimensions (le haut, le large et le lointain), le résultat de l'interférence
de ces trois tendances aura nécessairement un effet visible dans
l'espace : à tout moment on pourra l'analyser comme le résultat
d'un compromis entre la tendance à aller vers le bas, la tendance
à aller vers le haut, et la tendance à aller vers le lointain.
Si chacune de ces trois tendances
varie régulièrement, c'est-à-dire sans effectuer de
saut incompréhensible entre un instant et le suivant, le résultat
de l'interférence des trois tendances sera lui aussi nécessairement
régulier.
Certes, il pourra être compliqué,
très compliqué même, mais toujours on pourra le voir
dans l'espace, c'est-à-dire voir dans l'espace comment cela évolue
: si rien ne se passe c'est que les trois tendances se neutralisent, si
ça se déplace frénétiquement dans tous les
sens c'est que les trois tendances ne parviennent pas à trouver
un équilibre et ne cessent à tour de rôle de l'emporter
l'une sur l'autre, et si ça se déplace continuellement de
la même façon c'est que les trois tendances ne modifient que
très progressivement leur équilibre l'une par rapport à
l'autre.
Le problème
vient de ce qui se passe si une quatrième cause intervient en même
temps, car il manque une quatrième dimension à l'espace
pour que l'on puisse apprécier son influence en relation avec celle
des trois autres.
On dira alors que le phénomène
est déformé dans quatre dimensions simultanées, et
qu'il ne nous est visible que dans les trois dimensions de l'espace. Il
n'y aura plus coïncidence entre l'apparence dans l'espace et la réalité
des interactions,
parce que nous ne pourrons plus apprécier "de visu" l'influence
de l'une des causes d'interaction en même temps que l'influence des
trois autres. Ainsi, deux points pourront être très écartés
dans l'espace, tout en étant reliés directement l'un à
l'autre dans la 4ème dimension du phénomène.
3
tendances simultanées et indépendantes au déplacement
peuvent se décomposer en même temps sur chacune des 3 dimensions
de l'espace, mais 4 c'est trop !
Bien entendu, il ne faut pas voir
dans cette idée de 4ème dimension, la notion habituellement
attribuée par la science-fiction à une 4ème dimension
d'espace ou même d'espace-temps. Tout simplement, et ainsi qu'on
l'a dit, il s'agit seulement d'une 4ème cause influençant
le phénomène : deux points distants peuvent être soumis
à des "effets très proches" l'un de l'autre de la part de
cette 4ème cause, ce qui va les amener à faire des mouvements
similaires, de telle sorte qu'ils donneront l'impression de se déplacer
de concert malgré leur complète séparation dans l'espace.
Le chaos déterministe : dans la cellule aussi
Pour aider
à comprendre comment il est possible "qu'on ne puisse pas voir ce
qui se fait pourtant devant nos yeux", on peut donner l'exemple suivant
: si l'on dessine
un bonhomme sur une feuille de papier en deux dimensions (sa largeur et
sa hauteur), quelqu'un placé face à la feuille voit sans
problème ce qui se passe, alors que quelqu'un qui ne voit que sa tranche
(une seule dimension, celle de la hauteur de la feuille) est incapable
de voir ce qui se fait pourtant devant ses yeux. Il lui manque une dimension
pour voir, et donc pour comprendre ce qui se passe.
Depuis
le côté, il nous manque une dimension pour voir et
comprendre ce qui se passe
De la même façon, quand
un phénomène est produit par l'interférence de quatre
causes distinctes (phénomène dont nous dirons donc qu'il
est à quatre dimensions), l'espace possède un nombre insuffisant
de dimensions pour nous permettre de voir simultanément l'effet
de l'influence de ces quatre causes.
De façon générale,
les scientifiques ont pris l'habitude d'appeler "chaos déterministe"
ce type de situation où tout semble se produire de façon
incohérente selon ce qu'on en voit, alors que tout résulte
pourtant d'effets parfaitement déterministes d'après ce qu'on
en sait. Nous proposons que ce type de situation soit précisément
et seulement dû au débordement du nombre des dimensions de
l'espace par le nombre des dimensions qui interviennent dans le phénomène
et qui le déterminent.
Pour revenir aux chromosomes,
nous supposons donc ici que leur courant électromagnétique
est soumis à quatre tendances simultanées, et que l'absence
de forme visiblement cohérente pour son écoulement dans l'espace
et pour les tourbillons de protéines qui s'y dessinent en creux,
n'est due qu'à cette circonstance particulière qui nous empêche
de les appréhender, mais qui ne les empêche pas d'exister
et de fonctionner selon les principes que l'on a décrits.
1-
la première de
ces quatre tendances est l'entraînement même du courant qui
parcourt l'hélice du chromosome, et elle dépend de la séquence
des bases assemblées puisque chacune correspond à une courbure
différente du courant.
2-
la seconde tendance provient
de la présence d'autres chromosomes au voisinage : ils s'influencent
réciproquement, se modifient réciproquement la forme de leur
courant.
3-
la troisième tendance
provient du fait que chaque chromosome participe à un circuit bouclé
qui passe par le réseau des tubules de l'enveloppe de la cellule,
et qui revient vers le "centromère" du chromosome en empruntant
la canalisation du "centrosome" de la cellule. Ce cycle fermé équilibre
le débit du courant du chromosome, et il agit donc sur lui d'une
façon distincte des deux autres tendances qui tendent elles à
le déformer.
4-
la quatrième tendance
enfin, serait due à la présence de paires de chromosomes
appariés, l'un fonctionnant dans un sens, et l'autre dans l'autre
sens. Cette situation que l'on trouve dans les cellules "diploïdes"
(toutes nos cellules à l'exception des cellules sexuelles) tendrait
à équilibrer par paires et plus seulement en circuit fermé
individuel, l'ensemble des flux électromagnétiques qui circulent
dans les chromosomes. Pour cette raison elle conditionnerait donc d'une
4ème façon, non réductible avec les précédentes,
la forme du courant électromagnétique d'un chromosome.
Les protéines
ne seraient donc pas le résultat d'un code génétique
au fonctionnement compliqué et mystérieux, mais la simple
réplication de leur moule laissé en creux dans la forme des
chromosomes.
Le seul
problème, pour nous, serait que cette forme est comme en quatre
dimensions, alors que nous ne pouvons la voir qu'en trois dimensions seulement
dans l'espace.
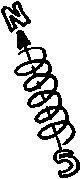 La
forme en hélice des chromosomes serait liée à leur
faculté de conduire un courant électrique et de se transformer
ainsi en aimant.
La
forme en hélice des chromosomes serait liée à leur
faculté de conduire un courant électrique et de se transformer
ainsi en aimant.
 Construction
du réseau des tubules d'une cellule pour recycler en permanence
le courant électromagnétique parcourant un groupe de chromosomes
Construction
du réseau des tubules d'une cellule pour recycler en permanence
le courant électromagnétique parcourant un groupe de chromosomes La
présence d'une protéine déforme le courant électrique
qui parcourt l'hélice des chromosomes et marque ainsi son empreinte
sur eux
La
présence d'une protéine déforme le courant électrique
qui parcourt l'hélice des chromosomes et marque ainsi son empreinte
sur eux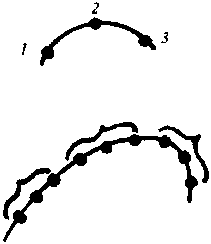 Par
3 points on ne peut faire passer qu'une seule courbe.
Par
3 points on ne peut faire passer qu'une seule courbe.
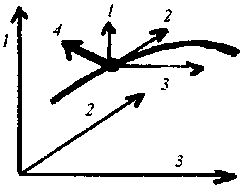 3
tendances simultanées et indépendantes au déplacement
peuvent se décomposer en même temps sur chacune des 3 dimensions
de l'espace, mais 4 c'est trop !
3
tendances simultanées et indépendantes au déplacement
peuvent se décomposer en même temps sur chacune des 3 dimensions
de l'espace, mais 4 c'est trop !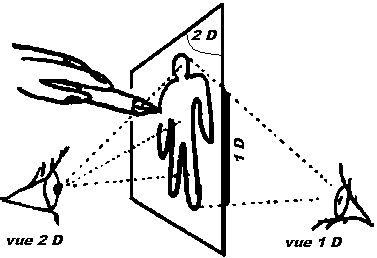 Depuis
le côté, il nous manque une dimension pour voir et
comprendre ce qui se passe
Depuis
le côté, il nous manque une dimension pour voir et
comprendre ce qui se passe