 La
présence d'une protéine déforme le courant électrique qui parcourt l'hélice des chromosomes et marque ainsi son empreinte sur eux La
présence d'une protéine déforme le courant électrique qui parcourt l'hélice des chromosomes et marque ainsi son empreinte sur eux |
| I
accueil |
E
sommaire Science |
avant
: E
chaos dans la cellule |
suite
: F
l'évolution génétique |
La duplication des protéines
On vient voir que, la plupart du temps, dans l'espace 3D, les chromosomes n'ont pas la forme de bâtonnets condensés qu'ils ont au moment de la division cellulaire, mais qu'ils n'en continuent pas moins à fonctionner sous l'influence conjointe des quatre principes d'organisation que l'on a listés au chapitre précédent. Comme on l'a dit, quatre effets distincts qui interfèrent entre eux, c'est quatre dimensions de fonctionnement, et c'est un de trop pour que nous puissions tous les voir en continu dans l'espace qui ne dispose que de trois dimensions.
On fait donc l'hypothèse que, dans leur état courant, les chromosomes, même s'ils perdent apparemment la forme de chromatide en hélice qui leur donne, au moment de la division cellulaire, l'aspect de bâtonnets bien organisés dans l'espace, n'en continuent pas moins à fonctionner comme s'ils étaient toujours sous cette forme de bâtonnet, même s'ils nous apparaissent alors sous forme de chromatine déroulée.
Cette hypothèse nous est indispensable puisque nous abordons
maintenant le fonctionnement des chromosomes en dehors des périodes
où la cellule se divise.
En dehors de la division cellulaire, les chromosomes fabriquent des protéines.
On peut supposer que c'est d'abord le hasard qui fait venir des molécules
de protéines, et qui les fait s'emmêler dans les chromosomes.
Mais on peut aussi supposer que ces molécules ne sont pas nées
au hasard, qu'il s'agit en fait de tourbillons qui sont nés entre
les chromosomes et qui ont migré progressivement vers eux.
Quelle que soit la cause exacte de sa venue, nous
allons maintenant suivre la façon dont l'une des protéines, ainsi restée prisonnière, va être dupliquée
par le chromosome où elle s'est incrustée.
| La
présence d'une protéine déforme le courant électrique qui parcourt l'hélice des chromosomes et marque ainsi son empreinte sur eux |
 |
 |
 |
Si la courbure du courant générée par chaque paire
de base est différente de celle générée par
l'autre paire, la courbure imposée par la protéine peut donc
se décomposer en petites séquences de courbes que 3 bases
groupées en séquence suffiront à définir précisément.
C'est
l'hypothèse que nous faisons sur la cause de la décomposition
du code génétique en séquence de 3 bases :
3 bases pour reproduire chaque portion minimum de la courbure du champ
magnétique engendré par la protéine.
Dans un autre texte [voir F
ce texte]
on rappelle comment, lorsque l'ADN est déroulé, les séquences
de base qui correspondent à une protéine, se trouvent dispersées
en des points parfois éloignés, séparés par
des séquences de base sans signification "codante" et que l'on appelle
"les introns".
Lorsque le courant magnétique de l'ADN est suffisamment fort
pour commencer à circuler de façon indépendante dans
chacune de ses 2 hélices, il déchire en divers endroits les
liaisons "hydrogène" qui collent les deux hélices, et chacune
reconstitue sa réplique.
Ces déchirures ne vont pas se faire au hasard, mais aux endroits
où vient se fixer un ARN que les généticiens appellent
"ARN Polymérase". On sait que cet ARN a la particularité
de faire tourner l'ADN à toute vitesse sur son axe. On fait ici l'hypothèse
que cette rotation a pour conséquence d'activer la circulation magnétique
à cet endroit, et donc de précipiter la duplication d'une
protéine particulière.
Sous l'impulsion de l'ARN polymérase, l'ADN commence donc à
répliquer le tourbillon de la protéine incrustée.
Comme ce tourbillon n'est pas encore reformé au début de
sa duplication, l'ADN
copie d'abord l'ensemble des séquences codant la protéine
et des séquences formant introns.
Cette première copie, les généticiens l'appellent
le "transcrit
primaire".
Dès que ce ruban transcrit est formé dans son entier,
le tourbillon de protéine auquel il correspond "prend" : les parties
"codantes" se mettent en continu et se séparent des introns.
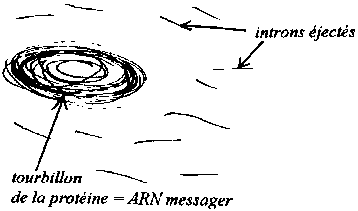 notre
hypothèse sur le processus d'épissage qui transforme le "transcrit
primaire" en "ARN messager" qui ne conserve que la partie "codante" du
transcrit primaire
|
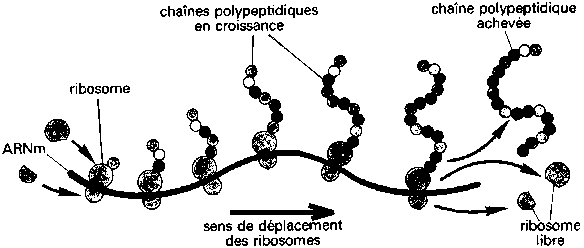
Le point intéressant est ce qui se passe dans la 2ème
sous-unité du ribosome. C'est là qu'est choisi le "bon" acide
aminé, celui qui propose le complément exact de la courbure
magnétique de chaque séquence de bases de l'ARN messager.
Dans cette sous-unité, c'est un autre ARN, que les généticiens
appellent "l'ARN de transfert", qui effectue ce choix.
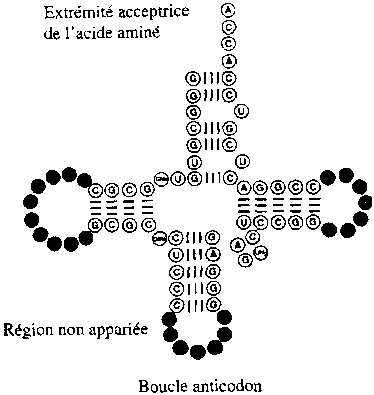 schéma
d'un ARN de transfert
[reproduction
d'un dessin de "l'Origine de la Vie"
par F.
Leroy aux éditions Biocosmos Centre] |
L'hypothèse
que nous formulons, est que
cette disposition de 4 boucles orientées dans l'espace, est destinée
à recréer
un équivalent en "négatif" d'un
fragment de la courbure magnétique exacte de la protéine
dans la situation où elle s'est incrustée dans le chromosome
:
- la boucle anticodon correspond à ce fragment
de courbure telle qu'elle était sur l'hélice d'ADN,
- et les deux autres boucles "neutres" auraient
pour fonction de recréer dans l'espace l'effet des 2 autres niveaux
d'enroulement de cette hélice encore en hélice, puis encore
en hélice.
|
Ainsi, toutes les particularités de la transcription de l'ADN, depuis la décomposition du code génétique en triplés de bases, jusqu'à la structure de l'ARN de transfert qui permet la traduction de ce code, vont dans le sens de notre hypothèse sur la nature "3 D" du code génétique. Cette hypothèse est que le code génétique ne fonctionnerait pas comme une suite linéaire d'instructions que les protéines lisent comme les ordinateurs lisent un programme informatique, il serait la réduction sous une forme linéaire (1 D) d'une suite d'empreintes magnétiques correspondant en réalité à une forme dans l'espace 3 D, et cette forme serait tout simplement la forme géométrique précise du courant magnétique produit par la protéine et déformant le courant magnétique propre des chromosomes dans l'espace à 3 dimensions. Ces 3 dimensions que recréent : - les triplés de bases du "code génétique", - puis les 3 boucles de l'ARN de transfert. |
Rappel : une version de ce texte, revue et améliorée dans le détail, est disponible en format pdf à l'adresse : http://www.quatuor.org/apparition-de-la-vie.pdf
| I
accueil |
E
Science |
G
haut |
suite : F l'évolution génétique |
|