AIRE TEGMENTALE VENTRALE
L'aire tegmentale ventrale (ATV) est constituée par un groupe de neurones localisés dans le mésencéphale. Ces neurones, qui contiennent de la dopamine, se projettent vers de nombreuses régions du cerveau en deux grandes voies : le système mésocortical et mésolimbique.
L'ATV fait partie du système de récompense. Elle est impliquée dans la motivation et certains processus cognitifs. Elle est également impliquée dans des pathologies comme l'addiction aux drogues et certains troubles psychiatriques.
AIRES DE BRODMANN
Le cerveau est organisé de manière très précise et très spécialisée. Plus exactement, il est organisé en zone, ce que Brodmann appelle des aires, dont voici le détail en terme de fonction :
Les aires de Brodmann 1, 2 et 3 : Les aires de Brodmann 3, 1 et 2(nommées dans cet ordre par convention), se confondent avec le gyrus latéral post central du lobe pariétal. Ce gyrus et ces aires sont classiquement associés à la fonction somatosensorielle primaire. Néanmoins, Kaas suggère que seule l'aire 3 de Brodmann correspond dans son organisation aux autres aires somatosensorielles : elle reçoit en effet la majeure partie des projections thalamiques couvrant les champs sensoriels (notamment, sensibilité cutanée superficielle), tandis que les aires 1 et 2 en reçoivent, en proportion, très peu (par exemple, sensibilité cutanée profonde pour l'aire 2).
L'aire de Brodmann 4 : On la fait classiquement correspondre à la fonction motrice primaire.
L'aire de Brodmann 5 : Elle est impliquée dans letraitement des informations somatosensorielles, il s'agit d'uncortex d'association.
L'aire de Brodmann 6 : L'aire de Brodmann 6 ou BA6 se situe sur le cortex frontal, en avant ducortex moteur primaire (aire de brodmann 4). Il se compose de deux aires fonctionnelles que sont lecortex prémoteur et l'aire motrice supplémentaire. Cette aire est également appelée Aire 6 agranulaire du fait de sa spécificité architectonique : il lui manque la couche granulaire (couche IV). Cette vaste région du cortex frontal est censée jouer un rôle dans la planification de tâches complexes etmouvements coordonnés.
L'aire de Brodmann 7 : Cette aire correspond à un cortex somatosensoriel associatif. Situé à la conjonction d'aires perceptives (en avant) et visuelles (en arrière), elle est censée jouer un rôle dans la coordination visuo-perceptive, intégrant les informations proprioceptives et visuelles, par exemple, pour déterminer la position relative d'un objet dans l'espace par rapport au corps ou à des parties de celui-ci (localisation dans l'espace). Il est donc impliqué, par exemple, dans les tâches de préhension, visuo-motrices (poursuivre visuellement un objet pour le saisir).
L'aire de Brodmann 8 : elle comprend notamment l'aire de coordination oculo-motrice. L'aire 8 joue un rôle dans laplanification de mouvement complexes. Sa partie basse, aire oculomotrice, joue un rôle dans lecontrôle des mouvements des yeux. On évoque l'aire 8 également dans lagestion de l'incertitude : une étude par imagerie par résonance magnétique suggère que l'activation de cette aire augmente avec le degré d'incertitude ressentie par l'individu.
L'aire de Brodmann 9 : Elle fait partie de la grande structure que l'on nomme cortex préfrontal dorsolatéral, impliqué dans de nombreuses fonctions telles que la planification, la mémoire de travail, la régulation de l'action et des fonctions intellectuelles supérieures.
L'aire de Brodmann 10 : L'aire 10 de Brodmann se situe dans le lobe préfrontal antérieur, dans une région connue comme la région frontale polaire, impliquée dans lesfonctions exécutives et la mémoire(récupération).
L'aire de Brodmann 11 : Fonction à déterminer. L’aire 11 constitue ce que l’on appelle le « cerveau social » avec d’autres aires cérébrales comme le gyrus cingulaire antérieur (ou aire 24 de Brodmann), l’amygdale (aire 34) et le gyrus temporal supérieur (aire 22).
L'aire de Brodmann 12 : L'aire 12 est particulièrement impliquée dans la mémoire des traits et des caractéristiques des objets.
L'aire de Brodmann 13 : aucun information complémentaire(seulement chez les primates).
L'aire de Brodmann 14 : Certaines hypothèses suggèrent que cette aire 14 est un cortex associatif lié au sens de l'olfaction ainsi qu'aux informations viscérales, jouant ainsi un rôle dans l'intégration des signaux du système nerveux autonome. Aucun information complémentaire(seulement chez les primates).
L'aire de Brodmann 15 : Cette aire a été définie chez le singe par Brodmann, mais il ne trouva pas de structure correspondante chez l'homme. Cependant, des données d'imagerie récentes suggèrent qu'une structure fonctionnelle homologue existe dans le cerveau humain.
L'aire de Brodmann 16 : Pas de données (à rechercher)
L'aire de Brodmann 17 : On la nomme également cortex strié. C'est l'une des plus anciennes structures du neocortex, spécialisée dans le traitement visuel des informations statiques et mobiles(mouvement) ainsi que dans lareconnaissance visuelle. On estime à 140 millions (Leuba et Kraftsik, 1994) le nombre de neurones que contient le cortex visuel primaire humain à l'âge adulte.
L'aire de Brodmann 18 : Elle a également reçu le nom decortex parastrié. Elle occupe lecuneus, le gyrus lingual et le gyrus occipital latéral.
L'aire de Brodmann 19 : Elle correspond au cortex extrastrié. Cette aire est considérée comme une aire associative (cortex visuel associatif), remplissant des fonctions visuelles telles que l'extraction d'un objet visuel, lareconnaissance de forme, l'attention visuelle ou l'intégration sensorielle multimodale.
L'aire de Brodmann 20 : Cette zone est connue pour jouer un rôle dans la reconnaissance visuelle(mémoire) et d'autres traitements visuels de haut niveau. Elle est également nommée région temporale inférieure.
L'aire de Brodmann 21 : Elle correspondant approximativement au gyrus temporal moyen. Cette région est connue pour jouer un rôle dans le traitement auditif, notamment des sons de la langue. Chez la plupart des individus, cette fonction est fortement latéralisée à gauche.
L'aire de Brodmann 22 : Le consensus actuel estime que cette région intervient dans le traitement auditif et la compréhension du langage. Cette fonction étant fortement latéralisée, les chercheurs pensent que l'aire 22 gauche intervient (chez les sujets dont l'hémisphère dominant est le gauche) dans la compréhension du langage (sémantique), tandis que l'aire 22 droite serait plutôt orientée vers le traitement de la mélodie, du rythme ou de l'intensité des sons parlés (prosodie).
L'aire de Brodmann 23 : Cette région est aussi connue sous le nom de cortex cingulaire postéro-ventral.
L'aire de Brodmann 24 : Francis Crick, l'un des découvreurs de l'ADN (1953), estime que cette zone est le siège de la volonté et du "libre-arbitre" : la lésion de cette aire entraine une aboulie et des difficultés motivationnelles. D'autres expériences semblent indiquer que cette zone intervient dans les réactions physiologiques émotionnelles (hérissement des poils, modifications cardiovasculaires, dilatation des pupilles). Son excitation peut par exemple provoquer un arrêt respiratoire et l'inhibition de la motricité volontaire.
L'aire de Brodmann 25 : L'aire 25 de Brodmann est une partie du cortex cingulaire constituée en bande étroite, dans le bas-avant du genou du corps calleux, d'où son nom de Subgenualis Area. Elle est liée à la région frontale 11.
L'aire de Brodmann 26 : Chez l'homme, cette région est une partie du cortex rétrosplénialappelée cortex ectosplénial. Le cortex est mince et les couches étroites. Fonction à déterminer.
L'aire de Brodmann 27 : Cette structure est impliqué dans letraitement olfactif, plus précisément dans la perception des odeurs, d'où l'appellation parfois de Cortex olfactif.
L'aire de Brodmann 28 : Associée à l'aire 34 (cortex entorhinal dorsal), elle constitue la majeure partie du cortex entorhinal. Fonction à déterminer.
L'aire de Brodmann 29 : Fonction à déterminer.
L'aire de Brodmann 30 : Fonction à déterminer.
L'aire de Brodmann 31 : L'aire 31 de Brodmann est la partie supérieure de l'arrière du cortex cingulaire, d'où son nom de Cortex cingulaire postérieur dorsal. Fonction à déterminer.
L'aire de Brodmann 32 : Cette région est associée aux fonctions de contrôle mental volontaire et à la pensée rationnelle, particulièrement mis en évidence par des tâches d'interférence type Stroop.
L'aire de Brodmann 33 : Fonction à déterminer.
L'aire de Brodmann 34 : Avec l'aire 28 (cortex entorhinal postérieur), elle forme le cortex entorhinal, lequel joue un rôle important dans les processus de la mémoire.
L'aire de Brodmann 35 : Avec l'aire 36, elle forme le cortex perirhinal. Ce cortex joue un rôle dans le mécanisme de récompense, et donc, dans le plaisir et la mémoire.
L'aire de Brodmann 36 : Fonction à déterminer.
L'aire de Brodmann 37 : Fonction à déterminer.
L'aire de Brodmann 38 : L'aire 38 de Brodmann comprend lesparties temporales polaires(antérieures) des gyri temporaux moyen et supérieur. Pour cette raison, on la nomme généralementPôle temporal.
L'aire de Brodmann 39 : Cette aire joue un rôle dans lacompréhension du langage, elle est fortement latéralisée et anatomiquement distincte d'un hémisphère à l'autre. Luria la considère comme une partie d'une zone temporo-pariéto-occipitale qui comprend l'aire 40, l'aire 19 et l'aire 37.
L'aire de Brodmann 40 : L'aire 40 de Brodmann se situe sur le cortex pariétal et constitue un point de jonction entre les lobes pariétal et temporal.
L'aire de Brodmann 41 : Avec l'aire 42, elle forme les cortex auditifs primaire et associatif.
L'aire de Brodmann 42 : Avec l'aire 41, elle forme les cortex auditifs primaire et associatif.
L'aire de Brodmann 43 : Cette aire est connue pour représenter le cortex gustatif.
L'aire de Brodmann 44 : Elle correspond à une partie du gyrus frontal inférieur, nommée Pars opercularis. Avec l'aire 45, elle forme l'aire de Broca. Les fonctions associées à cette zone sont fortement latéralisées : chez 95.5% des droitiers et 61.4% des gauchers, les aires 44 et 45 sont impliquées dans des fonctions sémantiques (notamment production du langage, hémisphère gauche). Certaines études suggèrent que l'aire 44 est davantage impliquée dans le traitement phonologique et syntaxique.
L'aire de Brodmann 45 : Fonction à déterminer.
L'aire de Brodmann 46 : Fonction à déterminer.
L'aire de Brodmann 47 : Fonction à déterminer.
L'aire de Brodmann 48 : Fonction à déterminer.
L'aire de Brodmann 49 : Fonction à déterminer.
L'aire de Brodmann 50 : Fonction à déterminer.
L'aire de Brodmann 51 : Fonction à déterminer.
L'aire de Brodmann 52 : Fonction à déterminer.
AMOUR - SENTIMENT AMOUREUX - DÉSIR
« Personne n’avait encore rapproché ces deux sentiments pour observer leur profil d’activation dans le cerveau, explique Jim Pfaus, professeur de psychologie à l’Université Concordia et coauteur de l’étude. Nous ne savions pas à quoi nous attendre – les sentiments auraient très bien pu être complètement distincts. Or, il s’avère que l’amour et le désir activent des zones précises mais apparentées du cerveau. »
Avec des confrères des États-Unis et de Suisse, le professeur Pfaus a analysé les résultats de 20 études consacrées à l’activité cérébrale observée chez des sujets auxquels on avait présenté des photos érotiques ou des clichés de leur partenaire. En fusionnant ces données, les chercheurs ont pu dresser une carte complète des circuits du sentiment amoureux et du désir dans le cerveau.
Ils ont ainsi découvert que deux structures cérébrales, l'insula (ou cortex insulaire) et le striatum, permettent de suivre la transformation du désir sexuel en sentiment amoureux. L’insula est une partie du cortex cérébral, repliée dans les profondeurs d’une zone située entre les lobes temporal et frontal, alors que le striatum siège à proximité, dans le prosencéphale.
L’amour et le désir sexuel activent différentes zones du striatum. Ainsi, la région touchée par le désir sexuel est généralement stimulée par des éléments éminemment agréables, comme l’acte sexuel et la nourriture. En revanche, celle qui est activée par le sentiment amoureux déclenche un processus de conditionnement qui confère une valeur intrinsèque aux stimuli procurant récompense ou plaisir. Autrement dit, à mesure que le désir sexuel se transforme en sentiment amoureux, ces éléments sont traités dans une zone différente du striatum.
Contre toute attente, cette zone du striatum correspond également à la partie du cerveau qui déclenche l’accoutumance aux drogues. Et pour cause, selon Jim Pfaus : « Le sentiment amoureux est une habitude qui dérive du désir sexuel à mesure que celui-ci est récompensé. Il opère de la même manière dans le cerveau que la toxicomanie. »
Bien que le sentiment amoureux puisse être une habitude, celle-ci n’est pas nécessairement mauvaise en soi. Dans le cerveau, l’amour active différentes voies qui exercent un effet déterminant sur la monogamie et l’attachement. Certaines zones cérébrales sont de fait moins actives lorsqu’une personne éprouve un sentiment amoureux que lorsqu’elle ressent du désir. « Alors que le désir sexuel a un objectif bien précis, l’amour est plus abstrait et plus complexe, et il est donc moins tributaire de la présence physique de l’autre », explique le professeur Pfaus.
Selon lui, les neurosciences cognitives ont permis aux chercheurs de définir précisément le siège de l’intelligence et de la résolution de problèmes dans le cerveau, mais il reste encore beaucoup à faire pour découvrir le centre précis du sentiment amoureux.
AMYGDALES
L'amygdale ou complexe amygdalien est un noyau pair situé dans la région antéro-interne du lobe temporal au sein de l'uncus, en avant de l'hippocampe et sous le cortex péri-amygdalien. Elle fait partie du système limbique et est impliquée dans la reconnaissance et l'évaluation de la valence émotionnelle des stimuli sensoriels, dans l'apprentissage associatif et dans les réponses comportementales et végétatives associées en particulier dans la peur et l'anxiété . L'amygdale serait également impliquée dans la détection du plaisir.
Structure en forme d’amande située près de l’hippocampe, l’amygdale joue un rôle essentiel dans la gestion de nos émotions et en particuliers nos réactions de peur et d’anxiété. Siège de nos émotions les plus primitives, l’amygdale reçoit des afférences directes de différentes modalités sensorielles et se projette sur de nombreuses régions cérébrales, comme l'hippocampe ou l’hypothalamus. L’action amygdalienne a donc un rôle de survie car c’est elle qui nous fait réagir en une fraction de seconde à la suite d’un stimulus menaçant. L’amygdale a également un rôle important dans la reconnaissance des émotions. Les patients victimes de lésions bilatérales de l’amygdale souffrent d’une véritable anesthésie des émotions et ne connaissent plus ni l’angoisse ni le plaisir.
L'amie de l'attention par Jean-Philippe LACHAUX
L'amygdale intéresse les chercheurs en neurosciences cognitives depuis longtemps à cause de son rôle dans l'apprentissage de la peur (pensez : « L'amygdale réagit aux mygales » – c'est un bon moyen mnémotechnique). Elle réagit aussi aux serpents. Un singe sans amygdales que l'on place devant une boîte transparente contenant de la nourriture et un serpent en plastique prend la nourriture sans hésitation. Un animal dont le cerveau est intact manifeste au contraire un comportement de peur et de frustration, et n'ouvre pas la boîte. Dans le même ordre d'idées, les patients souffrant de lésions cérébrales au niveau de l'amygdale ont du mal à reconnaître si un visage exprime ou non la peur.
Malgré sa petite taille, l'amygdale possède une capacité étonnante à reconnaître et mémoriser la valeur émotionnelle de ce qui nous arrive. Vous connaissez peut-être la célèbre expérience au cours de laquelle le physiologiste russe Ivan Pavlov parvint à entraîner un chien à saliver au simple son d'une cloche. Si un chien entend régulièrement sonner une cloche juste avant son repas, son cerveau finit par associer ce son à l'arrivée imminente de la nourriture, au point de réagir au son comme s'il s'agissait de la nourriture elle-même, en salivant. C'est ce que l'on appelle l'apprentissage par conditionnement, dont la découverte valut le prix Nobel à Pavlov en 1904.
Daniel Salzman et son équipe, à New York, se sont inspirés de Pavlov et ont montré à des singes des images en leur donnant un peu de nourriture après certaines d'entre elles et un petit jet d'air sur le nez, assez désagréable, après les autres. À force de conditionnement, les animaux finirent naturellement par développer un comportement attestant d'une préférence nette pour les images du premier groupe, de bien meilleur augure que les autres. Mais l'objectif des chercheurs n'était pas de traumatiser les singes, mais d'étudier le comportement de l'amygdale pendant l'apprentissage, et les résultats furent sans équivoque. Les neurones de l'amygdale, indifférents aux images au début de l'expérience, se mirent rapidement à réagir de façon sélective à l'un des deux groupes d'images. Certains neurones se mirent à réagir aux images « de bon augure », et d'autres aux images « de mauvais augure ». Ces neurones avaient repéré et mémorisé le caractère agréable ou désagréable de l'événement associé à chaque image. L'amygdale retient donc la valence, positive ou négative, d'un événement sensoriel, et cet apprentissage est flexible : à l'issue de l'expérience, Salzman et ses collègues changèrent la règle du jeu en donnant la nourriture après les images jusque-là « mauvaises » et le jet d'air après les images jusqu'ici « bonnes ». Les neurones, qui réagissaient aux images anciennement bonnes, ajustèrent leur réaction en quelques coups seulement pour ne plus réagir qu'aux autres images, et réciproquement, de façon à ce que chaque neurone continue d'annoncer soit le stimulus de bon augure, soit celui de mauvais augure, selon son ancienne préférence. L'amygdale est donc capable d'apprendre, de mémoriser et de reconnaître, rapidement et de façon flexible, le caractère positif ou négatif d'un stimulus. Grâce à l'amygdale, le cerveau peut donc étiqueter les événements qui parviennent à nos sens en fonction de leur caractère positif ou négatif.
L'amygdale sait reconnaître le caractère dangereux du serpent, certes, mais le fait-elle rapidement, et peut-elle ensuite amplifier l'activité du cortex visuel ? Concernant le deuxième point, la réponse est oui : l'amygdale est connectée directement à toutes les principales régions du cortex visuel, depuis le cortex visuel primaire jusqu'aux aires visuelles de plus haut niveau comme le gyrus fusiforme. Elle peut donc amplifier l'activité des neurones du cortex visuel et moduler la saillance de ce que nous voyons. La réponse au premier point paraît moins évidente ; l'amygdale n'est pas à proprement parler une aire visuelle, comment imaginer alors qu'elle puisse détecter la présence d'un serpent en moins de deux dixièmes de seconde ? Et pourtant, c'est possible, car l'information visuelle arrive au cortex par deux voies. La voie principale est celle dont nous avons déjà parlé, qui relie le cortex visuel primaire V1, à V2, V4 et ainsi de suite. C'est la « voie royale », chargée d'analyser toute la richesse du monde visuel.
Mais l'amygdale d'un patient dont le cortex visuel a été détruit réagit encore aux images émotionnelles, grâce à une seconde voie reliant la rétine à l'amygdale sans passer par les aires visuelles classiques. C'est une voie annexe capable d'informer l'amygdale de la présence d'éléments négatifs ou positifs dans l'environnement. Attention, toutefois : l'image n'est pas analysée de façon fine, mais seulement dans ses « grandes lignes ». C'est ce qui explique que nous puissions sursauter en prenant une corde pour un serpent. La voie visuelle amygdalienne n'est donc pas un double de la voie visuelle principale, et les patients qui n'ont plus de cortex visuel évoluent dans le monde quasiment en aveugle. Néanmoins, tous les éléments sont réunis pour faire de l'amygdale un centre important du mécanisme d'amplification de la réponse neuronale aux stimuli émotionnels, notamment visuels. Et quand l'amygdale ne fonctionne plus, ou mal, l'expérience montre que cette amplification n'a plus lieu. Ce mécanisme ingénieux s'applique aussi aux sons. En permettant au stimulus de générer une réponse neuronale plus forte, l'amygdale augmente automatiquement sa saillance et sa capacité à capturer l'attention. Il s'ensuit généralement une réorientation top-down de l'attention vers ce stimulus, dont l'importance est alors validée ou invalidée. Si le stimulus est effectivement nocif, ou positif, l'amygdale continue de lui attribuer la même étiquette ; sinon, l'amygdale s'adapte et change d'étiquette.
Et les choses ne s'arrêtent pas là. L'amygdale facilite également la mémorisation du stimulus et du contexte dans lequel il est apparu. La position de l'amygdale dans le lobe temporal médial la place au voisinage immédiat de plusieurs structures très importantes pour la mémoire, parmi lesquelles l'hippocampe, le cortex entorhinal et le cortex périrhinal, qui réagissent si bien à Marylin Monroe. Ces trois structures sont particulièrement importantes pour la mémoire épisodique — la mémoire des événements passés : « Hier, il faisait beau quand j'ai pris le bus. » Lorsque l'amygdale rentre en action, la communication entre ces régions est facilitée et l'événement est mieux mémorisé, ce qui explique, selon certains chercheurs, pourquoi nous nous souvenons si bien de ce qui nous a plu ou déplu, et pourquoi beaucoup se souviennent du lieu où ils étaient lorsqu'ils ont appris la nouvelle des attentats du 11 septembre 2001. L'amygdale est donc au centre d'un système efficace qui améliore la détection et la mémorisation des événements (potentiellement) les plus importants.
Amygdale droite (18/9/2011)
ANGOISSE (voir aussi Anxiété, Peur , Stress et SIA)
Henri Laborit : Parmi les fonctions du système nerveux central, on a peut-être trop privilégié ce qu’il est convenu d’appeler, chez l’homme, la pensée et ses sources, les sensations, et pas suffisamment apprécié l’importance de l’action, sans laquelle les deux autres ne peuvent s’organiser. Un individu n’existe pas en dehors de son environnement matériel et humain et il paraît absurde d’envisager séparément l’individu et l’environnement, sans préciser les mécanismes de fonctionnement du système qui leur permet de réagir l’un sur l’autre, le système nerveux. Quelle que soit la complexité que celui-ci a atteinte au cours de l’évolution, sa seule finalité est de permettre l’action, celle-ci assurant en retour la protection de l’homéostasie (Cannon), de la constance des conditions de vie dans le milieu intérieur (Claude Bernard), du plaisir (Freud). C’est lorsque l’action qui doit en résulter s’avère impossible que le système inhibiteur de l’action est mis en jeu et, en conséquence, la libération de noradrénaline, d’ACTH et de glucocorticoïdes avec leurs incidences vasomotrices, cardio-vasculaires et métaboliques, périphériques et centrales. Alors naît l’angoisse.
la corticosurrénale sécrète des glucocorticoïdes sous l’action d’un facteur hypophysaire, dit ACTH (hormone adrénocorticotrope), lui-même libéré par l’hypophyse, sous l’action d’un facteur hypothalamique, le CRF (corticotropin releasing factor), Or, celui-ci est libéré dans deux situations comportementales : la première, c’est lorsque le PVS est mis en jeu et que la fuite ou la lutte sont nécessaires pour conserver la structure vivante et la seconde dans une autre situation, lorsque le système inhibiteur de l’action est mis en jeu, Mais dans le premier cas, l’ACTH libérée, avant même de provoquer la sécrétion des glucocorticoïdes, agira sur l’activité du système nerveux en augmentant son incidence sur le fonctionnement moteur, L’ACTH va donc faciliter la fuite ou la lutte, Elle participe au fonctionnement de ce que nous avons appelé le système activateur de l’action (SAA) dont fait également partie le système de la récompense,
Si la fuite ou la lutte, nous l’avons vu, sont efficaces, les glucocorticoïdes vont stimuler le système inhibiteur de l’action qui mettra fin à l’action, laquelle action était efficace.
Les ennuis ne commencent que lorsque l’action s’avère inefficace, car alors le système inhibiteur de l’action va provoquer l’apparition d’une rétroaction positive en tendance, autrement dit d’un cercle vicieux. Ce système inhibiteur de l’action commandant par cascades successives la libération de glucocorticoïdes, ce qui ne peut encore que le stimuler. On ne peut donc sortir de ce cercle vicieux que par l’action dite «gratifiante», celle qui permet de rétablir l’équilibre interne et de fuir la punition. Il peut paraître curieux qu’après avoir insisté sur le fait qu’un système nerveux ne sert qu’à agir, nous signalions la présence dans l’organisation de ce système d’un ensemble de voies et d’aires aboutissant à l’inhibition de l’activité motrice. Cependant, ce système est malgré tout adaptatif, car dans certaines situations, mieux vaut ne pas réagir qu’être détruit par un agresseur mieux armé. L’ennui est que, si ce système d’évitement, permettant la conservation momentanée de la structure, n’est pas immédiatement efficace, si sa stimulation se prolonge, les remaniements biologiques résultant de son fonctionnement vont être à l’origine de toute la pathologie.
En effet, il existe un glucocorticoïde que tout le monde connaît, c’est l’hydrocortisone. Elle est utilisée en thérapeutique dans des cas bien précis, qui entrent généralement dans le cadre de ce que l’on appelle les maladies auto-immunes. Il s’agit d’affections dans lesquelles le système immunitaire n’est plus capable de reconnaître les propres protéines de l’organisme dans lequel il fonctionne et cette ignorance lui fait détruire des éléments parfaitement utiles et dont la disparition va être à l’origine d’affections diverses, le plus souvent chroniques, parmi lesquelles les arthroses sont l’exemple le plus courant. Mais les glucocorticoïdes sont extrêmement dangereux par ailleurs; en effet, tout médecin qui prescrit de la cortisone sait bien qu’il doit en même temps prescrire des antibiotiques. Pourquoi ? Parce que les glucocorticoïdes détruisent le thymus, glande qui est à l’origine de la libération des lymphocytes T, et favorisent la destruction ou l’inhibition d’autres cellules indispensables à l’activité immunitaire. Avec un système immunitaire déficient, sous l’action des glucocorticoïdes, l’organisme devient extrêmement fragile à l’égard de toutes les infections. De même, prennent naissance dans notre organisme des cellules non conformes, cellules néoplasiques qu’un système immunitaire efficace détruira au fur et à mesure de leur formation. Un système immunitaire inefficace en permettra la prolifération et autorisera donc l’évolution d’un cancer. Ainsi, on ne fait pas une maladie infectieuse et on n’est pas atteint d’une maladie tumorale au hasard, et la sécrétion par les surrénales d’une quantité démesurée de glucocorticoïdes fragilisera l’organisme dont la défense immunitaire se trouve paralysée.
Ce n’est pas tout. Les glucocorticoïdes, comme les minéralo-glucocorticoïdes retiennent aussi de l’eau et des sels. La masse des liquides extracellulaires va donc augmenter, tout comme la masse sanguine. Mais nous avons pu montrer que le système inhibiteur de l’action libérait également, à la terminaison des fibres sympathiques innervant les vaisseaux de l’organisme, de la noradrénaline. Celle-ci possède la propriété de provoquer une diminution du calibre (vasoconstriction) de tous les vaisseaux. Dans un système circulatoire à la capacité diminuée, une masse sanguine accrue va se trouver à l’étroit; il en résultera une pression supérieure à la surface interne de celui-ci. Il s’agit d’une hypertension artérielle, avec ses conséquences multiples telles qu’hémorragie cérébrale, infarctus viscéraux, infarctus myocardiques.
Il y a là, à notre avis, une autre différence avec la mise en jeu du système de la punition (PVS), qui, lorsque l’action est efficace, entraîne une mobilisation de l’organisme dans l’espace. Sa mise en jeu s’accompagne d’une libération d’adrénaline. L’adrénaline, à la différence de la noradrénaline, ne provoque une vasoconstriction qu’au niveau des vaisseaux cutanés et des vaisseaux de l’abdomen, réservant ainsi une masse de sang plus importante pour l’alimentation et l’évacuation des déchets des organes ayant, dans la fuite et la lutte, à fournir un travail supplémentaire: les muscles squelettiques, les vaisseaux pulmonaires, le cœur et le cerveau, ces derniers devant assurer l’approvisionnement d’un organe qui va permettre la mise en alerte, l’appréciation du danger et la stratégie à lui opposer. C’est la neurohormone de la peur, qui aboutit à l’action, fuite ou agressivité défensive, alors que la noradrénaline est celle de l’attente en tension, l’angoisse, résultant de l’impossibilité de contrôler activement l’environnement.
Les glucocorticoïdes vont aussi provoquer ce que l’on appelle un catabolisme protéique, c’est-à-dire détruire les protéines, éléments fondamentaux des structures vivantes. Le sommeil s’accompagne d’une restructuration protéique neuronale, les neurones au cours de leur activité ayant évolué vers un certain désordre moléculaire, qu’il s’agit de faire disparaître. Ainsi, en inhibition de l’action, le sommeil réparateur sera rendu plus difficile. On s’est aperçu d’ailleurs que l’injection d’un glucocorticoïde supprime le sommeil paradoxal chez l’animal. En inhibition de l’action, dans l’attente en tension, l’individu se trouvera donc insomniaque et fatigué. Depuis quelques années, on a pu mettre en évidence dans la majorité des états dépressifs une concentration élevée, anormale des glucocorticoïdes sanguins.
Par ailleurs, du fait de mécanismes complexes sur lesquels nous ne pouvons pas insister, on sait que les glucocorticoïdes participent également à l’apparition d’ulcères à l’estomac et d’autres affections dites «psychosomatiques» et qu’il serait préférable d’appeler d’«inhibition comportementale». Enfin, si l’angoissé «attend en tension» avec l’espoir encore de pouvoir agir, le déprimé, lui, paraît avoir perdu cet espoir.
ANXIÉTÉ (voir aussi Peur , Stress et SIA)
L'anxiété est une peur irrationnelle, sans objet, disproportionnée. L'angoisse désigne les sensations physiques qui accompagne l'anxiété (état psychique). L'anxiété peut être liée ou non à une représentation mentale, on dit qu'elle libre (flottante) ou liée. L'angoisse flottante est présente dans la
(qui est en fait considérée comme une " pseudo-névrose ", la bouffée d'angoisse signe l'absence d'une structure névrotique suffisamment élaborée mais il faut aussi savoir qu'elle apparaît en général dans d'autres pathologie). L'angoisse liée intervient dans le cadre d'une névrose plus structurée (quoique les mécanismes de défenses, bien que plus élaborés, ne suffisent pas à éviter le débordement émotionnel) : c'est (Freud). Enfin, lorsque l'angoisse présente des attaches psychiques à certains objets ou situation il s'agit d'une (actuellement appelée phobie).
Plusieurs observations montrent que l'excès de noradrénaline provoque l'anxiété. C'est la raison pour laquelle plusieurs médicaments anxiolytiques (c'est-à-dire contre l'anxiété) sont basés sur l'inhibition des neurotransmetteurs à noradrénaline, comme par exemple l'Avlocardyl ou le Propanolol.
Mais il existe aussi des médicaments qui, au contraire, vont amplifier le nombre de certains neurotransmetteurs « inhibiteurs », tels que les neurotransmetteurs « Gaba » : en effet, ceux-ci sont inhibiteurs parce qu'ils vont gêner la transmission de la noradrénaline. Parmi ces médicaments, vous connaissez certainement le Valium et le Lexomil.
AVC (accident vasculaire cérébral)
Chaque année en France, 130 000 personnes sont victimes d’un accident vasculaire cérébral et près de 50 000 en meurent.
L’accident vasculaire correspond à un arrêt brutal de la circulation sanguine au niveau du cerveau. Les AVC sont la plupart du temps dus soit à un caillot dans l’artère (et c’est le cas trois fois sur quatre), soit à une hémorragie, sorte de rupture des parois du vaisseau.
Les accidents ischémiques transitoires (AIT) ou éclipses cérébrales
Causes et facteurs de risque
Les signes de la maladie
BARRIÈRE HÉMATO-ENCÉPHALIQUE
Le cerveau est protégé par une barrière de l'invasion de substances étrangères transportées par le sang : la barrière hémato-encéphalique. Il n'est pas d'organe plus vascularisé que le cerveau ; pas une cellule nerveuse qui se trouve éloignée de plus d'un demi-centième de millimètre d'un capillaire sanguin. Mais ces capillaires cérébraux sont très différents de ceux qui irriguent le reste du corps. La paroi de ces derniers est faite d'une couche de cellules dites endothéliales, disjointes, ménageant entre elles des fenêtres par où s'écoulent les liquides et substances en solution. Le plasma sanguin, hormis la présence de grosses molécules de protéines, ne diffère pas du milieu extracellulaire ; les variations de composition de l'un reflètent les variations de l'autre. Les cellules endothéliales des capillaires cérébraux sont au contraire étroitement scellées entre elles, ne laissant aucun passage aux liquides et substances qui y sont dissoutes. Pour franchir la paroi, il faut traverser les cellules elles-mêmes, soit deux membranes plasmiques, l'une du côté de la lumière du vaisseau, l'autre du côté du cerveau. Barrière donc, mais barrière semi-perméable comme le sont les membranes plasmiques, c'est-à-dire laissant entrer et sortir l'eau et les solutés en fonction de règles physiques précises, selon des gradients ordonnés et par des modes de transport différents selon la nature des solutés. En poussant la comparaison, on peut assimiler le cerveau à une vaste cellule ceinte d'une double membrane ; au-dedans se trouve le milieu cérébral dans lequel baignent les cellules nerveuses, au-dehors, le milieu intérieur qui concerne le reste du corps.
Le passage à travers cette barrière est assuré par des systèmes de transport qui varient selon les substances. Les électrolytes, par exemple, qui confèrent aux neurones leurs propriétés électriques, obéissent à des règles strictes. Le moindre changement de leur concentration dans le milieu cérébral modifierait dangereusement l'excitabilité des cellules nerveuses.
La barrière hémato-encéphalique permet une véritable homéostasie du contenu ionique du cerveau. Il existe également des transporteurs pour les sucres, et l'on sait l'importance du glucose, carburant unique du neurone, pour les précurseurs des acides nucléiques, pour la choline, précurseur de l'acétylcholine, et pour certains acides aminés précurseurs des neurotransmetteurs et des neurohormones. En l'absence de transporteur, il n'y a pas de passage. C'est le cas pour la dopamine, d'où l'échec de son utilisation comme médicament pour suppléer dans la maladie de Parkinson à son manque dans le cerveau. Le précurseur, la L. DOPA, passe par contre la barrière, d'où son efficacité thérapeutique. A ce propos, nous ne ferons qu'évoquer le problème posé par l'existence de la barrière, lors de l'utilisation de médicaments destinés au cerveau. Des substances, actives si elles sont administrées directement dans le cerveau, se révèlent bien souvent inefficaces lorsqu'on les injecte par voie sanguine. La recherche pharmaceutique s'efforce de trouver un produit actif dont la conformation chimique permette la traversée de la barrière : c'est un mime, molécule différente mais douée des mêmes affinités et effets que la molécule imitée.
Comme toute réalité gênante, la barrière hémato-encéphalique suscite le doute et flirte parfois avec le mythe. On a évoqué la possibilité d'un transport à l'intérieur de vésicules formées à partir des membranes. L'utilisation d'un marqueur, la peroxydase du radis noir, enzyme protéique dont le trajet peut être révélé et suivi au microscope, a confirmé l'existence de la barrière : dans un sens comme dans l'autre, l'enzyme ne traverse pas. Il en est de même pour les peptides et protéines. Les hormones ne peuvent ni pénétrer ni sortir du cerveau, à l'exception toutefois des stéroïdes, solubles dans la membrane. Le système de communication hormonal en milieu cérébral n'est donc pas celui du milieu intérieur. Il existe toutefois des portes de sortie pour les hormones du cerveau ; elles sont étroitement circonscrites à quelques territoires spécialisés où la barrière n'existe pas.
Grâce à son mur et à des systèmes de transport spécialisés, voici constitué le milieu autonome qui entoure la cellule nerveuse. Il importe avant tout que ce milieu reste constant et qu'il assiste, impassible, aux événements qui agitent le milieu intérieur. De même que ce dernier avait permis l'indépendance de l'organisme vis-à-vis de l'environnement, le milieu cérébral permet l'indépendance du cerveau vis-à-vis du milieu intérieur. Cette homéostasie du milieu cérébral doit être soumise aux mêmes restrictions que celle du milieu intérieur : elle est parcellaire et variable dans le temps.
CAPTEURS (OU RÉCEPTEURS) SENSORIELS EXTÉROCEPTIFS
Ils sont au nombre de 5 :
1 - LA PEAU
Le cerveau reçoit les informations en provenance du revêtement cutané par deux voies :
2 - la voie réticulée (reticulus = petit réseau) lente et plus diffuse. Elle est faite d'un cablage en réseau. La collecte des messages se fait par la moëlle épinière et ils sont ensuite dirigés vers le tronc cérébral où ils s'attardent dans le fouillis inextricable de la substance réticulée avant de gagner le thalamus et éventuellement le cortex cérébral. (d'ou la sensation douloureuse ou thermique difficile à localiser correctement et la persistance lorsque le stimulus a cessé.
Ainsi la connectivité mélange des informations de provenances et qualités différentes.
Chaque partie sensitive du corps est représentée proportionnellement à sa richesse en terminaisons sensitives (notamment : mains, lèvres, langue, pieds)
Le sens du toucher est en fait le regroupement de plusieurs sens : la faculté de détecter le froid, la chaleur, la douleur et la pression. Si vous comparez l'étendue des aires du cerveau dévolues à chacun des sens, vous verrez que le centre du toucher est significativement plus grand que les zones réservées à l'odorat et au goût.
2 - LA LANGUE
Tout d'abord pour être "senti" l'aliment doit être mélangé avec la salive car les papilles gustatives ne fonctionnent qu'en milieu liquide ,elles captent donc l'aliment . Viens ensuite l'élaboration du signal qui sera transmis au cerveau par des protéines. Ce dernier analyse et restitue les sensations grâce aux minuscules récepteurs sensoriels situés dans chaque papille. Les terminaisons nerveuses de ces récepteurs sont en effet reliées au cerveau qui va analyser ces sensations et les mémoriser.
Le signal est ensuite amener dans l'aire gustative située dans le buion, c'est-à-dire en plaisir. On parle de " mémoire gustative ".
3 - LE NEZ
C'est le regroupement de tous les axones olfactifs qui forment le nerf olfactif (nerf n° I). Une partie du nerf olfactif est en relation avec l'hypothalamus qui joue un rôle important dans les comportements primitifs comme la prise d'aliments ou le comportement sexuel.
Depuis les doux arômes de la fleur à la puanteur des poubelles, le cerveau de quelqu'un qui a un « blair » particulièrement sensible - par exemple un chef cuisinier ou un oenologue - peut interpréter 10 000 odeurs différentes tandis que le commun des mortels doit se contenter de 3 000. Le nez des gens ne varie pas beaucoup en sensibilité; toute la différence réside dans l'aptitude d'une personne à se concentrer sur les odeurs, à les identifier et à se souvenir de celles qui lui sont familières. Le sens de l'odorat est aussi une capacité d'adaptation nécessaire à la survie. La possibilité de reconnaître une viande avariée par l'odorat, par exemple, nous protège contre des aliments dangereux.
4 - LES YEUX
Une vie haute en couleurs
Chassé-croisé
Partie de chasse
Deux yeux valent mieux qu'un
Dans le brouillard
Voir, c'est croire ?
5 - LES OREILLES
L'oreille se divise en trois parties : l'oreille externe, l'oreille moyenne et l'oreille interne. La partie externe est la plus visible, mais la moins importante. Ce n'est qu'un petit bout de peau et de cartilage (le pavillon) sur chaque côté de la tête, dont la fonction principale semble surtout de nous rendre ridicules ! L'oreille externe reçoit les ondes sonores et les dirige dans le conduit auditif. Ce dernier, qui mesure 2,5 cm, les achemine à l'intérieur de la tête où elles frappent le fond, c'est-à-dire le tympan (membrane tympanique pour les savants), fin morceau de peau de la taille de l'ongle du petit doigt. Les changements de
pression de l'air font vibrer le tympan.
De bonnes vibrations
Baissez le volume, s'il vous plaît
SYSTÈME VESTIBULAIRE
Le système vestibulaire informe le système nerveux central (tronc cérébral et cervelet) sur la position et les déplacements de la tête.
Le système vestibulaire contribue à la coordination des mouvements de la tête et du corps.
L'oreille interne, appelée également labyrinthe vu la complexité de sa forme, contient deux organes sensoriels aux fonctions différentes.
N.B. : La gentamicine, est un antibiotique connu pour léser les structures de l'oreille interne, au point d'occasionner éventuellement la surdité, des tintements d'oreilles et des dégâts considérables au système d'équilibration. Mais parce que la gentamicine est bon marché et efficace, on continue à la prescrire, en général pour une courte durée.
CATÉCHOLAMINES
Les catécholamines sont des composés organiques synthétisés à partir de la tyrosine et jouant le rôle d'hormone ou de neurotransmetteur.
Les catécholamines les plus courantes sont l'adrénaline (épinéphrine), la noradrénaline (norépinéphrine) et la dopamine. Elles sont synthétisées par les cellules de la médullo-surrénale et par les neurones postganglionnaires du système nerveux orthosympathique. L'adrénaline agit en tant que neurotransmetteur dans le système nerveux central et comme hormone dans la circulation sanguine. La noradrénaline est principalement un neurotransmetteur du système nerveux sympathique périphérique, mais se retrouve présente dans le sang.
L'état de stress augmente le taux de catécholamines dans le sang. Au cours d'une activité physique, les catécholamines induisent des modifications physiologiques de l'organisme : augmentation de la fréquence cardiaque, de la pression artérielle et du taux de glucose dans le sang.
CELLULE NEUROGLIALE
Les autres cellules du cerveau sont les cellules gliales, les méconnues, celles dont on oublie de parler. Neurones et cellules gliales sont comme maîtres et serviteurs. De par leur excitabilité, les premiers possèdent, comme nous l'avons vu, la propriété noble de conduire l'information sous forme d'un signal électrique et d'assurer la conversation entre neurones. Pour oubliées qu'elles soient, les cellules gliales sont pourtant dix fois plus nombreuses que les neurones. Mais elles sont inexcitables, dépourvues de synapses, et on ne leur attribuait jusqu'alors que des fonctions sans prestige : entretien, emballage, protection, nutrition, etc. Des traités consacrés au cerveau ignorent souvent les cellules gliales. Ce serait introduire l'inexcitable dans l'excitable, le mou et le gluant dans la dentelle précieuse tissée par les neurones !
Et pourtant, les cellules gliales sont inséparables des neurones. D'abord, par une origine commune chez l'embryon ; ensuite, par leur disposition anatomique. Elles occupent tout l'espace libre laissé par les neurones, s'infiltrant entre leurs corps cellulaires et enveloppant leurs prolongements. Ne laissant qu'un espace extracellulaire réduit à quelques millionièmes de millimètre, mais ininterrompu et permettant la libre circulation des solutés. Inséparables enfin, par leurs propriétés physiologiques, les cellules gliales n'exercent leurs fonctions qu'en liaison avec les neurones voisins. Alors que ces derniers sont individualisés et ne communiquent entre eux qu'au travers des espaces spécialisés, les synapses, les cellules gliales établissent entre elles des jonctions ou courts-circuits qui leur donnent une continuité et une sorte d'identité commune. La glie réalise des ensembles indiscrets autour d'unités neuronales discrètes.
La gaine membranaire formée par les cellules gliales autour des prolongements neuronaux contient une substance graisseuse, la myéline, qui donne sa coloration blanche à la matière cérébrale et aux nerfs. Interrompue à intervalles réguliers par les noeuds de Ranvier, cette gaine isolante force l'influx à sauter d'un noeud à l'autre, accélérant ainsi sa propagation. La glie conditionne donc par sa présence la vitesse de transit des signaux. La disparition de la myéline au cours de certaines affections ou son absence congénitale compromettent gravement les fonctions nerveuses, comme on peut l'observer sur une souche de souris mutantes, les Jimpys, chez lesquelles la myéline ne se développe pas normalement.
Le rôle sécréteur de la glie a été très clairement suggéré par Nageotte, une histologiste française du début du siècle, qui écrit : « La névroglie est une glande interstitielle annexée au système nerveux. » Les cellules gliales sécrètent en effet des neurotransmetteurs malgré l'absence de formation synaptique. Dennis et Miledi a ont montré qu'après section d'un nerf moteur les cellules gliales qui viennent occuper la place des fibres nerveuses dégénérées sont capables de libérer de l'acétylcholine spontanément ou en réponse à une stimulation électrique, comme l'auraient fait les neurones intacts. On insiste beaucoup, depuis quelques années, sur la capacité des cellules gliales à sécréter du GABA, substance qui, dans le cerveau, est le principal neurotransmetteur inhibiteur. Cette capacité de sécrétion est couplée avec celle de capter le GABA présent dans l'espace extracellulaire. C'est ce que l'on peut vérifier en impressionnant une émulsion photographique par les molécules de GABA radioactif captées par la cellule, technique dite de l'autoradiographie. On découvre ici une fonction possible de la glie : le stockage des neurotransmetteurs et leur libération dans certaines conditions en parallèle avec le jeu synaptique que mènent ces agents de liaison interneuronale.
On parle souvent de la fonction nutritive de la glie comme si celle-ci allait de soi et sans trop savoir ce qu'elle recouvre. Il est possible en effet que les cellules gliales captent et apportent aux neurones des précurseurs des neurotransmetteurs — une sorte de garde-manger. Il est plus
important, toutefois, d'insister sur la propriété qu'ont gardée les cellules gliales de se diviser, au contraire des cellules neuronales. Rakic et Sidman, dans une série d'études consacrées au développement du cortex cérébral et du cervelet chez le singe et l'homme, ont montré que les cellules gliales et leurs prolongements formaient une sorte de trame ou filet sur lequel les neurones migraient vers leur destination finale. La possibilité de se reproduire permet aux cellules gliales de jouer un rôle dans la réparation des lésions nerveuses en occupant l'espace perdu et en guidant et favorisant la pousse d'éléments nerveux de remplacement grâce à la sécrétion de facteurs de croissance. Une protéine qui favorise la croissance des cellules nerveuses (le NGF) peut être extraite des tumeurs gliales.
Nous nous attarderons sur les propriétés électriques des cellules gliales, au risque d'être paradoxal, étant donné l'absence d'excitabilité et de synapse de ces cellules. Comme celle des neurones, leur membrane est polarisée, la face interne étant négative par rapport à la face extracellulaire, et ce potentiel de membrane négatif, plus élevé que celui des neurones voisins, constituerait une sorte de puits de négativité attirant les charges positives voisines. Ce potentiel dépend principalement de la répartition d'un ion positif, le potassium, de part et d'autre de la membrane. Lorsque la concentration de potassium s'élève à l'extérieur, cet ion positif tend à pénétrer dans la cellule et à dépolariser la face interne négative de la membrane. Les variations lentes de potentiel que l'on enregistre avec une électrode à l'intérieur de la cellule traduisent fidèlement les mouvements de l'ion potassium à travers la membrane, eux-mêmes fonction de la concentration extracellulaire de l'ion. Or celle-ci dépend de l'activité électrique des neurones voisins. Un neurone qui produit des potentiels d'action crache du potassium. Lorsque le neurone bat de façon répétitive, le potassium s'accumule en dehors de la cellule. L'excitabilité neuronale étant fonction du potassium extracellulaire, plus le neurone décharge, plus il devient excitable. En pompant le potassium accumulé à l'extérieur du neurone, la cellule gliale résorbe l'excès d'ion et prévient les risques d'emballement.
Comme on vient de le voir, l'entrée du potassium dans les cellules gliales provoque des variations lentes de potentiels qui suivent comme une ombre l'activité des neurones voisins. Une expérience de Kelly et Van Essen démontre élégamment cette participation gliale à l'activité électrique du cerveau. Une électrode de verre est placée à l'intérieur d'une cellule gliale du cortex visuel d'un chat. On sait que certains neurones des aires corticales visuelles sont activés de façon spécifique par le déplacement d'une cible lumineuse dans une direction déterminée de l'espace. On observe ici que la cellule gliale présente des variations lentes de potentiel dont l'amplitude répond au déplacement de la cible lumineuse dans une direction donnée. Ces variations correspondent donc vraisemblablement à l'activation d'une colonne de neurones voisins. Si ces variations lentes n'entraînent pas la genèse d'influx, il n'est pas exclu, en revanche, qu'elles provoquent la libération de substances qui pourraient, en retour, modifier le fonctionnement des synapses voisines.
Il faut insister sur le caractère non spécifique d'une telle signalisation. Il s'agit d'un effet non circonscrit au niveau d'une synapse. La réponse gliale indique seulement le niveau de trafic dans les neurones voisins. L'existence de communication électrique entre les cellules gliales elles-mêmes augmente ce caractère de diffusion. Deux exemples nous permettront d'illustrer ces rôles possibles de la glie. Un premier exemple relève de la pathologie. Ce serait un fonctionnement anormal de la glie qui, en favorisant l'accumulation du potassium dans les régions du cerveau, provoquerait leur embrasement électrique et serait à l'origine d'une crise d'épilepsie.
(JD VINCENT) : Les cellules gliales occupent les espaces entre les neurones et forment un ensemble compact avec des espaces intercellulaires de quelques dizaines de nanomètres. Elles n'établissent entre elles aucun contact synaptique de type chimique, mais peuvent être jointes par des formations spéciales appelées jonctions gap et jonctions serrées.
On distingue cinq types de cellules gliales répartis dans le système nerveux central et périphérique.
Les cellules gliales du système nerveux périphérique sont dites cellules de Schwann. Je n'en parlerai pas, car elles servent d'isolant aux fibres du système nerveux périphérique, fonction dévolue dans le cerveau aux oligodendrocytes.
Les astrocytes sont de petites cellules (10 pm de diamètre) munies de nombreuses ramifications. On en distingue deux types : le type 1 entre en contact avec des neurones et des vaisseaux sanguins ; le type 2 n'entre en contact qu'avec des neurones.
L'ensemble des astrocytes joue un rôle de protection des neurones et de contrôle de la composition du milieu extracellulaire. Ils régulent en particulier la composition en potassium, en absorbant une partie des ions libérés par les axones lors du passage d'un potentiel d'action. De la même façon, les astrocytes participent à l'élimination des neurotransmetteurs libérés dans les fentes synaptiques, grâce à des systèmes protéiques de recapture.
Les oligodendrocytes, localisés au sein des faisceaux d'axones, forment une gaine de myéline autour de certains axones du système nerveux central. Leurs expansions constituent des languettes membranaires qui s'enroulent autour des axones. Ces segments myélinisés' sont séparés par des points non myélinisés : les noeuds de Ranvier.
Les cellules de la microglie sont peu nombreuses : elles ne représentent que 5 à 10 % des cellules gliales centrales. Il semble qu'elles jouent un rôle essentiel au cours du développement embryonnaire ; stade durant lequel elles se différencient à partir de monocytes sanguins qui ont franchi la barrière hémato-encéphalique. Elles forment alors la microglie amiboïde. Ces cellules se différencient ensuite en microglie ramifiée dont le rôle chez l'adulte reste encore très mal connu, en relation notamment avec les processus immunitaires au sein du cerveau.
Les cellules épendymaires forment un épithélium qui tapisse les parois des ventricules cérébraux et du canal de l'épendyme de la moelle épinière. Elles sont unies entre elles par des jonctions serrées qui assurent la cohésion de l'épithélium. Certaines d'entre elles présentent de nombreux cils qui baignent dans le liquide céphalo-rachidien. D'autres, les tanycytes, possèdent des microvillosités sur leur surface apicale et des prolongements qui viennent au contact des capillaires, des neurones ou d'autres cellules gliales.
CERVEAU
Le cerveau est le siège de toutes les commandes de l’organisme. Il est situé dans la boîte crânienne constituée par les os du crâne, soutenu par les méninges et ainsi protégé ; cette boîte est inextensible. Le cerveau comprend plusieurs régions : les deux hémisphères cérébraux, le cervelet, en relation les unes avec les autres et avec les organes ou d’autres parties du corps. Le tronc cérébral est sous le cervelet et met en communication le cerveau avec la moelle épinière qui se trouve dans la colonne vertébrale.
Le cerveau appartient au système nerveux central.
Fonction du cerveau
Le cerveau est ainsi responsable du rythme cardiaque et respiratoire, des fonctions qui pour nous sont inconscientes. Mais il est aussi impliqué dans la prise de décision, dans la motricité du corps, le comportement, la mémoire, la conscience…
Le cerveau a été cartographié et certaines zones semblent associées à des fonctions précises (aire de la parole…), alors que les fonctions plus complexes (la mémoire, la conscience) semblent être beaucoup plus réparties.
Structure du cerveau
Le cerveau est composé de deux hémisphères, droit et gauche, connectés entre eux par le corps calleux composé de substance blanche (axones gainés de myéline). Divisés en lobes, les hémisphères sont les parties les plus développées chez les mammifères.
Le cerveau est divisé en plusieurs parties, en fonction de leur origine embryonnaire :
Le cortex cérébral constitue la couche externe du cerveau et comporte la majorité des neurones (substance grise). Les méninges sont des enveloppes protectrices (dure-mère, arachnoïde et pie-mère) entre le cortex et l’os du crâne. Le cortex cérébral est subdivisé en cinq lobes cérébraux en fonction de leur position.
Le cerveau est l'organe de notre corps chargé de la perception et de l'interprétation du monde extérieur. Il est composé d'une myriade de cellules nerveuses, appelées neurones, qui forment un réseau de connexions extrêmement efficace.
Pour donner un ordre de grandeur, on estime que le cerveau contient un peu plus de 100 milliards de neurones dont chacun peut former jusqu'à 10.000 connexions, le tout contenu dans un volume équivalent à celui d'une brique de lait. C'est grâce à ce formidable réseau de neurones que nous pouvons entre autres apprécier un bon film, tomber amoureux ou résoudre une énigme.
Anatomie du cerveau
Cette différence de couleur provient du fait que le cortex contient principalement les corps cellulaires de neurones, alors que la substance blanche est constituée d'un agglomérat d'axones qui relie les différentes aires corticales les unes aux autres. Cette différence de couleur provient du fait que le cortex contient principalement les corps cellulaires de neurones, alors que la substance blanche est constituée d'un agglomérat d'axones qui relie les différentes aires corticales les unes aux autres.
Le cerveau est subdivisé en plusieurs parties, que l'on peut distinguer en se basant d'une part sur son développement et, d'autre part, sur son architecture.
Le cerveau est triplement protégé (de l'extérieur vers l'intérieur) :
Les hémisphères du cerveau
Pour se repérer, on définit un axe antérieur-postérieur qui va de la face vers l'arrière du cerveau. En dessous des hémisphères cérébraux, on distingue d'autres structures comme l'arrivée de la moelle épinière et le cervelet.
D'un point de vue évolutionniste, le pont et le bulbe rachidien font partie des structures les plus anciennes du cerveau. Ils sont responsables de fonctions majeures, indispensables à la survie, comme les battements cardiaques ou la respiration.
Formation du cerveau
C'est au cours du développement embryonnaire que se construit le cerveau : 250 000 neurones se créent chaque seconde chez le fœtus ! Mais dès 25 ans, le nombre de neurones baisse inexorablement à raison de 100 000 en moins par jour. Car contrairement aux autres cellules, les neurones ne se renouvellent pas. Mais pas de panique ; nous ne devenons pas pour autant plus bêtes, car le cerveau développe des circuits alternatifs pour compenser ces pertes.
Né à partir de l'ectoblaste (ecto=en-dehors), feuillet qui enveloppe l'embryon, il passe successivement par les phases suivantes:
Le tube neural va donner la totalité du système nerveux central et c'est le renflement de son extrémité antérieure qui donnera le cerveau.
Les entrées et les sorties :
Il n'y a que deux entrées possibles dans le cerveau, la nerveuse et l'humorale. Cette dernière est parfaitement contrôlée au niveau de la barrière hématoencéphalique. L'entrée nerveuse laisse parvenir au cerveau des données recueillies et mises en forme au niveau des organes des sens et de récepteurs spécialisés où déjà s'ébauche la représentation, qu'il s'agisse du monde extérieur (extérocepteurs), de la position du sujet dans le monde (propriocepteurs) ou du milieu intérieur (intérocepteurs).
Deux sorties également : la nerveuse, qui permet la réalisation des programmes moteurs, et l'humorale, qui se fait sous forme de libération d'hormones au niveau d'une région spécialisée ou carrefour hypothalamo-hypophysaire. Ces sorties hormonales, comme les sorties motrices, peuvent se faire en réponse à des stimulations venues du corps et de l'environnement ou selon des programmes centraux. Parmi ceux-ci, certains sont réglés par des horloges logées dans le cerveau. Ce sont des montres molles à la Dali, elles s'allongent et se raccourcissent au gré des humeurs et de l'environnement.
Professions travaillant sur le cerveau :
CERVELET
Le cervelet est un centre nerveux régulateur de la fonction motrice, au sens large (mouvement + posture + équilibre). Il reçoit des informations de tous les segments du névraxe (moelle épinière, tronc cérébral, cerveau). Il traite ces informations pour donner, aux programmes moteurs du mouvement, une organisation chronologique et somatotopique (organisation temporo-spatiale). Il assure ainsi la régulation :
Situation du cervelet
Les cellules de Purkinje sont des neurones GABAergiques du cortex cérébelleux, découvertes par Jan Purkinje au cours du XIXe siècle. Ce sont les cellules nerveuses les plus complexes et elles interagissent avec des centaines de milliers d'autres cellules nerveuses, créant ainsi plus de connexions qu'aucune autre cellule du cerveau. Elles envoient leurs axones vers les noyaux profonds du cervelet, pour parfois atteindre le thalamus, le mésencéphale et la moelle épinière. Les faisceaux sensoriels acheminant l'information de l'organisme vers le cervelet aboutissent généralement au niveau des cellules de Purkinje. Les noyaux profonds sont des stations relais pour des impulsions nerveuses spécifiques.
Comme le cerveau, le cervelet est divisé en deux hémisphères. Chaque hémisphère cérébelleux est composé de trois lobes. Le lobe flocculo-nodulaire est une zone qui reçoit, depuis les oreilles, les informations sensorielles nécessaires au maintien de l'équilibre. Le lobe antérieur reçoit, en provenance de la moelle épinière, les informations sur ce que fabrique le reste du corps. Enfin, le lobe postérieur communique avec le cerveau.
Les deux hémisphères du cervelet sont reliés par un faisceau étroit de fibres blanches appelé vermis. Le vermis a de tout temps été considéré comme important. Le médecin grec ancien Galien pensait qu'il contrôlait le pneuma (c'est-à-dire l'esprit vital). Cette idée était fausse, comme beaucoup des hypothèses de Galien, mais elle a été admise pendant des siècles. Les scientifiques savent aujourd'hui que le vermis joue le rôle de relais entre le cervelet et la moelle épinière.
Les amygdales cérébelleuses sont une autre partie du cervelet. Il existe deux amygdales cérébelleuses, chacune associée à un hémisphère. Elles n'ont pas une grande importance fonctionnelle, mais interviennent dans beaucoup de maladies du cervelet. Dans certains cas, l'hypertrophie des hémisphères cérébelleux (le gonflement du cerveau) va engager les amygdales dans le trou occipital. Cet état donne lieu à de nombreux symptômes.
Le cervelet a trois fonctions principales : le maintien de l'équilibre, la régulation du tonus musculaire et la coordination des mouvements. Il participe au maintien de l'équilibre grâce à sa relation avec le système vestibulaire, lequel est impliqué dans la coordination et l'équilibre. Tout commence avec l'influx en provenance de l'oreille interne qui voyage le long du nerf vestibulaire jusque dans le cerveau (le nerf vestibulaire est également appelé nerf crânien VIII. En même temps qu'il préserve la position normale du corps, au repos comme en mouvement, le cervelet participe aussi à la production du tonus musculaire nécessaire au maintien de cette position.
Même les plus balourds d'entre nous n'ont pas de difficultés à se tenir debout et à marcher sans se casser la figure. Nous n'accordons pas d'attention à ces actions, mais notre cervelet travaille à la vitesse de l'éclair pour assurer notre équilibre.
Le cervelet n'est le siège d'aucune activité consciente, il travaille automatiquement à coordonner un ou plusieurs muscles en mouvement. Souvent comparé à un ordinateur, il traite des informations en provenance des oreilles, des yeux, des muscles et des tendons, et, sans les stocker, il y répond immédiatement de façon à modifier la position des membres.
Comment se déroule ce processus ? L'organisme est doté d'une boucle de rétrocontrôle entre les systèmes musculaires et nerveux. Vous en apprendrez plus sur le rôle des nerfs dans ce rétrocontrôle au prochain chapitre. Pour l'instant, il est important de comprendre que les muscles, les tendons et les articulations disposent de détecteurs spéciaux, appelés « propriocepteurs », qui indiquent quand ces parties du corps s'étirent, se contractent et se penchent. Les messages concernant ce que font ces parties du corps sont envoyés au cervelet, qui n'initie pas le mouvement, mais analyse et ajuste la progression des mouvements.
Le cerveau évalue ce que font les muscles et détermine s'ils agissent en fonction de ses instructions. Il envoie alors de nouveaux ordres au cervelet afin de réaliser tout ajustement nécessaire. Le cervelet relaie les ordres au système musculaire. Ce processus est quasi instantané et doit être répété pour chaque mouvement des muscles.
Par exemple, quand vous essayez de renvoyer une balle de tennis, le cerveau décide en premier lieu de frapper la balle. Après avoir traité l'information visuelle (la balle arrivant vers vous) et l'information tactile (la raquette que vous tenez), le cerveau, par l'intermédiaire de ses connexions motrices, vous permet de frapper la balle. Le cervelet modifie votre geste afin que vous situiez la raquette à proximité de la balle.
L'activité cérébrale du serveur est plus compliquée. Le cervelet du serveur relaie aux muscles les instructions nécessaires pour déterminer la vitesse, l'effet et la direction de la balle. Dans le cas du serveur comme du receveur, vous pouvez constater que le cortex moteur et le cervelet doivent fonctionner ensemble pour réaliser les mouvements et les contrôler.
A l'inverse, marcher ne nécessite pas d'implication cérébrale. Le cervelet est l'agent principal intervenant pour modifier la marche. Sans l'influence du cervelet, vous ne tiendriez pas debout.
L'importance du cervelet est mise en évidence par l'observation des problèmes neuromusculaires induits par des maladies ou des lésions le concernant. Celles-ci provoquent des erreurs dans la vitesse, la portée, la force et la direction des mouvements volontaires, de même que des oscillations irrégulières dans le geste. Les personnes présentant des dysfonctionnements cérébelleux souffrent de tremblements, perdent l'équilibre et présentent des troubles de coordination. En bref, vous auriez du mal à
l'équilibre et toucher le bout de votre nez avec votre doigt sans votre « petit cerveau ».
CLAUSTRUM
Le claustrum (ou avant-mur) est une fine couche de matière grise cérébrale située entre le putamen en dedans et l'insula en dehors, dont il est séparé par les capsules externe et extrême, respectivement.
Le claustrum entretient des connexions réciproques avec le néocortex, ainsi qu'avec l'hypothalamus latéral, l'amygdale basol-latérale, le noyau basal de Meynert, le corps genouillé latéral et le noyau thalamique intralaminaire centro-latéral.
Il possède une carte rétinotopique et somatosensorielles.
Il pourrait jouer un rôle dans la conscience
COORDONNÉES DU CERVEAU
Les géographes du cerveau disposent d'un système de coordonnées qui porte le nom de TALAIRACH, l'un de ses créateurs. Plutot que dire "le foyer fusiforme gauche se trouve ici ou là par rapport à tel sillon" je peux dire qu'il se trouve, en moyenne aux coordonnées X = -45, Y = -60 et Z = -10, où ces 3 chiffres donnent la position dans les sens droite-gauche, avant-arrière et haut-bas, par rapport à un point de référence conventionnel.
COLLICULUS - COLLICULI
Les colliculi (du latin « colliculus » signifiant : mont, colline) ou tubercules quadrijumeaux sont des masses arrondies de l'encéphale humain.
Ils sont au nombre de quatre : deux antérieurs (les colliculi supérieurs), deux postérieurs (les colliculi inférieurs). Ils se situent entre le cervelet et le cerveau, au niveau de l'aqueduc de Sylvius (nouvellement rebaptisé aqueduc du mésencéphale). La glande pinéale ou épiphyse est située entre les tubercules postérieurs et antérieurs.
Le colliculus supérieur coordonne les mouvements des yeux et de la tête, tandis que le colliculus inférieur fait partie de la voie auditive ascendante entre l'oreille et le cortex auditif
Atonio Damasio :
Les couches profondes, quant à elles, contiennent, outre une carte du monde visuel, des cartes topographiques des informations auditives et somatiques, ces dernières en provenance de la moelle épinière ainsi que de l'hypothalamus. Les trois variétés de cartes - visuelles, auditives et somatiques - sont d'ordre spatial. Cela signi- fie qu'elles sont compilées avec une telle précision que les informations fournies par l'une, par exemple visuelle, cor- respondent à celles d'une autre carte liée à l'ouïe ou à
aspects des états du corps ne sont aussi superposées, au sens littéral, ce qui favorise leur intégration. Celle-ci est d'autant plus significative que ses résultats peuvent avoir accès au système moteur (via les structures toutes proches du gris périaqueducal, ainsi que via le cortex cérébral).
Qu'en est-il du collicule supérieur chez l'homme ? Sa destruction sélective est rare chez les êtres humains, si rare que la littérature neurologique n'en enregistre qu'un seul et unique cas, portant sur des dégâts bilatéraux, qui ont heureusement été étudiés par le grand neurologue et spécialiste de neurosciences Derek Denny-Brown15. La lésion était due à un traumatisme crânien et le patient a survécu plusieurs mois, dans un état de conscience gravement handicapée et proche du mutisme akinétique. Sa mentalisation était elle-même compromise. Je dois cependant ajouter que j'ai, en une occasion, rencontré un patient au collicule atteint, mais chez lequel on ne détectait qu'un bref trouble de conscience.
Voir avec les seuls collicules quand les cortex visuels sont atteints pourrait consister à sentir qu'un objet X non déterminé se déplace dans l'un des quadrants de la vision, c'est-à-dire s'écarte de moi ou s'approche. Dans aucun de ces deux cas, je ne pourrais décrire mentalement ce qu'est l'objet et il se peut que je n'en aie même pas conscience. Nous parlons ici d'un esprit très vague, rassemblant des informations partielles sur le monde, même si le fait que les images soient vagues et incomplètes ne les rend pas inutiles, comme le montre la vision aveugle. Toutefois, lorsque les cortex visuels font défaut à la naissance, comme dans le cas des hydroencéphales décrits plus haut, les collicules supérieurs et inférieurs peuvent avoir un apport substantiel au processus mental.
Il faut encore ajouter un autre fait attestant la contribution des collicules supérieurs à l'esprit. Le collicule supérieur produit des oscillations électriques de type gamma, phénomène qu'on a relié à l'activation synchronique des neurones. Le neurophysiologue Wolf Singer a suggéré que ce serait corrélé à la perception et même à la conscience. À ce jour, le collicule supérieur est la seule région du cerveau située hors du cortex cérébral dont on sait qu'elle produit des oscillations de type gamma.
CORPS GÉNICULÉ LATÉRAL
Le corps géniculé latéral (corpus geniculatum laterale, anciennement corps genouillé externe) du thalamus est une partie du cerveau qui traite l'information visuelle provenant de la rétine. Les termes scientifiques "géniculé" et "genouillé" viennent du latin geniculatum, geniculatus « courbé à la manière d'un genou plié ».
Le corps géniculé latéral (CGL) reçoit l'information directement de la rétine et envoie des projections dans le cortex visuel primaire, dans la couche 4c. Mais ce cortex visuel V1 exerce une forte rétroaction sur le CGL puisque les 8/10e des connexions excitatrices qui y pénètrent proviennent de V1 et de structures sous-corticales.
Les cellules ganglionnaires de la rétine envoient leurs longs axones au corps géniculé latéral via le nerf optique, le chiasma optique et le tractus optique. Ils sont plus d'un million de fibres à suivre cette voie qui contourne le mésencéphale, chemine sur la face médiale du lobe temporal et aboutit (pour 80% d'entre elles) dans le corps géniculé latéral.
CORPS STRIÉS
Les corps striés sont les centres de la motricité volontaire ; quand ils fonctionnent mal comme dans la maladie de Parkinson, les patients éprouvent de grandes difficultés à décider de leur mouvement et le font avec de grands tremblements.
chez les vertébrés supérieurs, on distinguera trois noyaux dans le corps strié : les noyaux «caudé», «putamen», et «pallidum».
CORTEX CÉRÉBRAL
Le cortex est une couche de 2 millimètres qui recouvre une surface équivalant à une serviette de plage (2 m2). Si bien que pour se loger dans une boîte crânienne qui ne fait qu'un litre et demi (une bouteille d'eau classique), le cortex fait des plis appelés circonvolutions ; ce sont ces circonvolutions qui permettent aux neurologues d'établir la cartographie du cortex. Car ces deux millimètres d'épaisseur, qui n'ont l'air de rien, renferment tout de même six étages de cellules, qui permettent l'abstraction. Le cortex renfermerait ainsi quelque vingt milliards de neurones et comme il n'est pas programmé à la naissance, c'est en fait notre disque dur, l'ensemble de nos mémoires.
Le cortex cérébral est, à l'instar de tous les cortex de l'organisme, la partie périphérique des hémisphères cérébraux. Le cortex cérébral est le siège des fonctions neurologiques élaborées. Il s'agit de l'intelligence, du mouvement volontaire, de la conscience, de la sensibilité etc.
Le cortex joue un rôle primordial en ce qui concerne les fonctions nerveuses de base. Il s'agit des différentes modalités que sont la motricité, la sensibilité, la sensorialité ou sensoricité. D'autre part le cortex tient également le rôle indispensable dans d'autres fonctions supérieures que sont le langage et la mémoire etc.
On a tendance à comparer le cortex cérébral à un ensemble de circuits électroniques bien qu'il soit plus complexe dans sa structure et son fonctionnement.
Chaque couche de neurones composant le cortex est constituée, quand on l'observe au microscope électronique, d'un réseau de corps cellulaires et de prolongements neuronaux.
Les frontières entre les lobes sont constituées de sillons, profonds et de scissures (scissures de Sylvius et de Rolando) sur la surface du cortex. La scissure de Sylvius est grosso modo horizontale dirigée d'avant en arrière sur la face latérale de chacun des hémisphères cérébraux et séparant les lobes frontal et pariétal du lobe temporal. La scissure de Rolando, appelée également scissure central, est un sillon profond du cortex cérébral, situé à la surface de chacun des hémisphères cérébraux et qui sépare le lobe frontal du lobe pariétal.
On distingue trois variétés ou trois types de cortex dont la complicité va croissante et qui se différencient par leur structure observable au microscope électronique ainsi que par leur rôle :
L'archicortex permettant la survie de l'organisme et les comportements élémentaires de celui-ci.
Le paléocortex comprenant l'hippocampe, siège de la mémoire, de l'attention, l'émotion et des comportements selon son expérience passée
Le néocortex constitué de petites zones qui permettent au cerveau d'organiser son fonctionnement. Ces zones sont appelées aire corticale primaire dont chacune est responsable de certains types d'activités.
Chaque hémisphère cérébrale comprend quatre lobes :
Le cortex cérébral est constitué d'aires d'association situées autour de toutes les aires primaires. Ces aires d'association ont la capacité essentiellement de permettre la coordination des fonctions de base. La perception et la compréhension des images et du son fait intervenir les aires d'association. D'autres part les aires d'association ou aire secondaires, situées au voisinage des précédentes traitent les informations qui sont fournies par celle-ci. Leur rôle est essentiel en ce qui concerne la reconnaissance des objets perçue par la voie sensorielle correspondante. Les agnosies qui sont les incapacités ou l'impossibilité de reconnaître, de façon générale, correspondent à des troubles d'intégration à ce niveau.
Les aires tertiaires pariéto-temporale et frontale reçoivent, quant à elles, des informations en provenance de plusieurs aires secondaires. Ces aires assurent les fonctions supérieures comme le langage, les fonctions intellectuelles, l'organisation dans l'espace et le contrôle du comportement.
Chaque hémisphère joue un rôle particulier. L'hémisphère gauche est le siège de la parole, des mouvements, de la sensibilité de la moitié gauche du corps. Il existe un phénomène de croisement c'est-à-dire que l'hémisphère droit assure certaines fonctions à gauche et inversement pour l'hémisphère gauche. Ceci est une règle générale sauf chez certains gauchers.
On parle de dominance hémisphérique chez les droitiers qui possèdent donc un hémisphère gauche dominant. Le langage, dans ce cas, est assuré par le fonctionnement de l'hémisphère dominant donc le gauche chez le droitier et généralement le droit chez le gaucher.
L'hémisphère droit quant à lui joue un rôle beaucoup plus important que ce que l'on a cru jusqu'ici. Il assure ainsi différentes fonctions essentielles comme la prosodie ou si on préfère la tonalité du discours, le contenu émotionnel et l'organisation du discours ainsi que la reconnaissance de l'humour et la compréhension des métaphores, du sens et des images des mots. Enfin l'hémisphère droit possède des capacités de récupération après une atteinte lésionnelle de l'hémisphère gauche.
CORTEX CINGULAIRE
Cette région ancienne du cortex est située sur de nombreuses voies remontant du cerveau émotionnel limbique inconscient, ainsi que sur de nombreuses autres voies descendantes conscientes et impliquées dans la planification de l'action en provenant du cortex préfrontal.
Il semblerait exister une baisse d'activité de cette région cérébrale lors d'une dépression nerveuse et à l'inverse une hyperactivité en cas de crise maniaque.
C'est la partie antérieure du cortex cingulaire qui est plus active lors d'une dépression, contrairement au reste de cette structure.
Cette structure cérébrale agirait un peu comme une interface entre les régions cérébrales inférieures instinctives et le néocortex dont les actions sont rationnelles
Chez le sujet normal l'activation de cette structure cérébrale contribuerait à focaliser notre attention en particulier en provenance de notre propre corps comme la douleur ou des sentiments négatifs lors de la dépression.
Le cortex cingulaire antérieur (CCA) est la partie frontale du cortex cingulaire, qui ressemble à un « collier » s'enroulant autour du corps calleux dont les fibres relaient les signaux neuronaux entre les hémisphères cérébraux droit et gauche.
Il comprend une zone ventrale et une zone dorsale et semble jouer un rôle dans une grande variété de fonctions autonomes comme la régulation de la pression artérielle et du rythme cardiaque, les fonctions cognitives, telles que l'anticipation de récompense, la prise de décision, l'empathie et l'émotion.
CORTEX ENTORHINAL
Le cortex entorhinal est un important centre de la mémoire dans le cerveau. Le cortex entorhinal constitue la principale entrée à l'hippocampe et est responsable du traitement préalable des signaux d'entrée. Dans le cadre de réponses « réflexes », l'association des impulsions de l'œil et l'oreille se produit dans le cortex entorhinal. Le système cortex entorhinal-hippocampe joue un rôle important dans la consolidation de la mémoire et de son optimisation durant le sommeil.
CORTEX FRONTAL
Le lobe frontal du cerveau est limité en arrière par la scissure de Rolando et par la scissure sylvienne (de Sylvius). Il comprend le tiers avant de l'hémisphère cérébral.
L'écorce motrice du lobe frontal c'est-à-dire la frontale ascendante, zone numéro 4 de Brodmann contient des cellules pyramidales géantes de Betz et les axones au départ de la voie pyramidale (corticospinale). Chaque partie de l'écorce motrice de la frontale ascendante permet de bouger une partie du corps. La partie interne et supérieure de la frontale ascendante exerce un contrôle sur les mouvements des membres inferieurs et du pied. La partie inférieure gère les mouvements du visage, du larynx et de la langue. L'écorce motrice des hémisphères droit gère les mouvements de la moitié gauche du corps et inversement.
L'écorce motrice correspondant à la zone 6 de Brodmann commande les actes complexes et ordonnés mais en cas de lésions de cette zone la mobilité volontaire est gardée.
Les centres moteurs du langage se trouvent au niveau de la zone 44 de Brodmann et dans les centres de Broca. Le centre moteur du langage se situe dans le tiers postérieur de F2 et F3 au niveau de l'hémisphère qui domine c'est-à-dire gauche chez le droitier. Ce centre contrôle la transmission des idées et le langage écrit et parlé. Une atteinte du cerveau à ce niveau entraîne une aphasie (altération du langage faisant suite à une lésion cérébrale) motrice.
CORTEX INFÉRO-TEMPORAL
Le cortex inféro-temporal est spécialisé dans la reconnaissance des objets. En théorie, il est possible de savoir ce qu'une personne regarde en mesurant l'activité de son cortex inféro-temporal. Chez le singe, les chercheurs ont trouvé des neurones qui réagissent fortement à la vue d'une banane, mais presque pas à la vue d'une pomme. Et lorsqu'un singe doit chercher une banane, ces neurones maintiennent un niveau d'activité élevé pendant tout le temps que dure la recherche. C'est un mécanisme semblable à la préchauffe de Desimone qui permet une réaction neuronale rapide dès qu'une banane apparaît dans le champ de vision de l'animal. Ce mécanisme sert aussi à la mémoire de travail : lorsqu'un singe doit mémoriser l'image d'une banane, les neurones qui réagissent à la vue de la banane maintiennent également une activité élevée pendant le temps où cette image est maintenue en mémoire, exactement comme si le singe était en train de chercher cette banane. Si le chercheur montre ensuite aux singes deux photos, représentant l'une une pomme et l'autre une banane, ce dernier sait indiquer sans difficulté le fruit qui lui a été présenté quelques instants auparavant. Donc pour ces neurones du lobe temporal, il n'y a pas de différence entre chercher un objet autour de soi et le garder en mémoire. Plusieurs études ont confirmé cette prédiction : en demandant à un singe de simplement mémoriser un dessin de banane et en lui montrant ensuite sans qu'il s'y attende plusieurs dessins de fruits, David Soto et son équipe ont constaté que l'animal orientait spontanément son attention et son regard vers la banane, comme si celle-ci était plus saillante que les autres fruits.
Cela signifie que notre attention est spontanément attirée par ce qui est relié à ce que nous avons en tête à un moment donné, ce qui occupe nos pensées. Ce phénomène a un corollaire intéressant. Si vous marchez dans la rue en vous fredonnant un air des Beatles, il y a de grandes chances qu'en passant devant la librairie, vous remarquiez Paul McCartney en couverture d'un des livres en vitrine, tout simplement parce que cette photo rappelle la chanson que vous avez en tête. La photo de Paul McCartney en couverture d'un livre est un exemple de stimulus rappelant les Beatles, tout comme un bout de la chanson « Yesterday » ou le nom « John Lennon », écrit sur un bout de papier. Ces trois stimuli n'ont rien en commun sur le plan purement physique, car l'un est un son et les deux autres des images très différentes l'une de l'autre ; et pourtant, il existe des neurones à l'avant du lobe temporal, qui réagissent indifféremment à tout ce qui fait penser aux Beatles'. En fait, le simple fait de penser au Beatles suffit à activer ces neurones ; ils sont sensibles aux concepts, perçus ou imaginés. Quand vous avez en tête une chanson des Beatles, ces neurones sont actifs, et sont donc dans un état de sensibilité accrue, prêts à réagir dès qu'un stimulus rappelant les Beatles se présente, et c'est pourquoi la photo de Paul McCartney en vitrine attire facilement votre attention.
Notre attention est donc spontanément attirée par ce qui nous préoccupe à un moment donné. Il peut s'agir d'une préoccupation à très court terme, comme la faim : lorsque nous avons faim, les images évoquant la nourriture attirent plus facilement notre attention'. Mais il peut aussi s'agir d'un intérêt plus stable, de votre passion du moment. Il est bien connu qu'au début de leur première grossesse, les jeunes femmes se mettent tout d'un coup à voir des ventres ronds partout. Le monde leur semble tout d'un coup différent, plein de femmes enceintes et de petits bébés. Cet effet est très utilisé en publicité : lorsqu'un événement de grande ampleur comme la coupe du monde de football occupe l'esprit de toute une population, les annonceurs attirent l'attention sur leurs produits en les enrobant d'un emballage rappelant l'événement en question. Pendant toute une période, les gens vivent dans un monde différent où tout ce qui rappelle le foot est plus visible. C'est pourquoi il existe un fromage officiel de la coupe du monde.
CORTEX INSULAIRE ou INSULA
Le cortex insulaire est situé au fond du sillon latéral. Sa surface est marquée par cinq gyrus de l'insula : trois gyri courts en avant et deux autres plus longs en arrière. Il est encerclé par le sillon circulaire qui marque la séparation avec les parties operculaires des lobes temporaux, frontaux et pariétaux.
Le cortex insulaire est divisé en deux parties : une large insula antérieure et une petite insula postérieure, dans laquelle ont été identifié plus d'une douzaine d'aires différentes. La partie antérieure du lobe de l'insula, une composante des aires prémotrices, joue un rôle dans la production du langage et sa partie postérieure contient des aires somatosensorielles secondaires intégrant des informations en provenance de différentes modalités.
Le cortex insulaire joue un rôle dans diverses fonctions, principalement liées aux émotions ou à la régulation de l'homéostasie du corps. Ces fonctions incluent la perception, le contrôle moteur, la conscience du soi, le fonctionnement cognitif et les expériences interpersonnelles. Cette partie du cerveau est donc fortement impliquée dans certains dysfonctionnements psychopathologiques.
L’insula est une région du cerveau longtemps restée méconnue à cause de sa position en profondeur dans les replis du cortex. Et comme elle n’était pas non plus associée aux fonctions dites « supérieures », cela la rendait moins intéressante aux yeux de ceux qui s’intéressaient à l’étude de la conscience.
Mais cette indifférence s’est transformée en un vif regain d’intérêt à la suite des travaux de Antonio Damasio et de son hypothèse des marqueurs somatiques qui fait la part belle à l’insula.
Damasio a en effet proposé que cette région du cortex permettrait de cartographier nos états viscéraux qui sont associés à des expériences émotionnelles, donnant ainsi naissance à un sentiment conscient. Ces travaux s’inscrivent dans le courant de la cognition incarnée où la pensée rationnelle consciente ne peut être séparée des émotions et de leur incarnation dans le reste du corps.
L’insula, aussi appelé cortex insulaire, produirait donc un contexte émotionnel adéquat pour une expérience sensorielle donnée. Elle est d’ailleurs bien située pour intégrer de l’information relative à l’état du corps et rendre cette information disponible pour des processus cognitifs et émotionnels d’ordre supérieur. L’insula reçoit par exemple des inputs sensoriels «homéostatiques» par l’entremise du thalamus, et elle envoie des outputs à plusieurs structures reliées au système limbique, comme l’amygdale, le striatum ventral et le cortex orbitofrontal.
L’insula est aussi déjà bien associée aux processus de douleur ainsi qu’à plusieurs émotions de base comme la colère, la peur, le dégoût, la joie ou la tristesse. Sa portion la plus antérieure est considérée comme faisant partie du système limbique. L’insula serait aussi grandement impliquée dans les désirs conscients, comme la recherche active de nourriture ou de drogue. Ce qu’il y a de commun dans tous ces états, c’est qu’ils affectent le corps entier en profondeur. Un constat qui tend à renforcer son rôle probable dans la représentation que nous nous faisons de notre propre corps ainsi que dans l’aspect subjectif de l’expérience émotionnelle.
Enfin, l'insula humaine, et à un moindre degré celle des grands singes, aurait deux innovations évolutives qui lui permettrait de porter la lecture de notre état corporel à un niveau inégalé chez les autres mammifères. D'abord la partie antérieure de l'insula, et plus particulièrement de l'insula de l'hémisphère droit, serait davantage développée chez les humains et les grands singes que chez les autres espèces animales. Ceci permettrait un décodage plus précis de nos états viscéraux, et donc par exemple à une simple mauvaise odeur de devenir un sentiment de dégoût, ou encore au toucher d'une personne aimée de se transformer en sentiment de délice.
L'autre modification majeure à notre insula est la présence d'un type de neurone que l'on retrouve seulement chez les grands singes et l'humain.
Il s’agit de grandes cellules nerveuses allongées en forme de cigare appelées VEN. De plus, on ne retrouve ce type de neurone que dans l’insula et le cortex cingulaire antérieur. Ces neurones font des connexions avec diverses parties du cerveau, ce qui serait un atout essentiel pour les fonctions supérieures qu’on attribue à ces deux structures cérébrales.
Par Antonio Damasio :
La partie arrière de l'insula se compose du néocortex moderne et sa partie médiane est d'un âge phylogénétique intermédiaire. On sait depuis longtemps que le cortex insulaire est associé au fonctionnement des viscères : il les représente et participe à leur contrôle. Avec les cortex somatosensoriels primaire et secondaire (SI et SII), l'insula produit des cartes corporelles. Par rapport aux viscères et au milieu intérieur, l'insula est l'équivalent des cortex visuel et auditif primaires.
À la fin des années 1980, j'ai formé l'hypothèse que les cortex somatosensoriels joueraient un rôle dans les sentiments et j'ai indiqué que l'insula pouvait en produire. Je voulais m'écarter de l'idée, me semblait-il sans espoir, consistant à attribuer l'origine des états de sentiment aux régions régulant l'action, comme l'amygdale. À l'époque, le fait de parler d'émotion suscitait une sympathie amusée et celui de suggérer que les sentiments avaient un substrat distinct créait un froid.
Depuis 2000, nous savons toutefois que l'activité de l'insula est un important corrélat de tous les types concevables de sentiments, de ceux qui sont associés aux émotions à ceux qui correspondent à toutes les formes de plaisir ou de douleur, induites par une large gamme de stimuli — entendre de la musique qu'on aime ou qu'on déteste ; voir des images qu'on aime, y compris érotiques ; être à court de drogue et se sentir en manque ; etc. L'idée selon laquelle le cortex insulaire est un substrat important des sentiments est certainement correcte.
L'insula n'est pourtant pas le fin mot de l'histoire quand il s'agit des corrélats des sentiments. Le cortex cingulaire antérieur devient actif en parallèle avec l'insula quand nous ressentons des sentiments. Ce sont des régions étroitement imbriquées, toutes les deux unies par des connexions mutuelles. L'insula a une double fonction sensorielle et motrice, quoiqu'elle soit plutôt orientée sur l'aspect sensoriel du processus, alors que le cortex cingulaire antérieur opère comme structure motrice.
Ce qui est le plus important, bien sûr, c'est le fait que plusieurs régions sous-corticales jouent un rôle dans la construction des états sentimentaux. Au premier abord, on pouvait considérer que des régions comme le nucleus tractus solitarius et le noyau parabrachial transportaient les signaux issus de l'intérieur du corps, qu'ils véhiculaient vers un secteur dédié du thalamus, lequel en retour les transmettait au cortex insulaire. Toutefois, comme indiqué plus haut, il est probable que les sentiments commencent à émerger à partir de l'activité de ces noyaux, étant donné leur statut spécial – ces noyaux sont les premiers réceptacles des infonnations venues des viscères et du milieu intérieur, dans la mesure où ils sont aptes à intégrer les signaux de tout l'intérieur du corps ; dans le chemin qui mène de la moelle épinière à l'encéphale, ces structures sont les premières à pouvoir intégrer les signaux de tout le paysage interne, poitrine et abdomen, viscères, ainsi que les aspects viscéraux des membres et de la tête.
Dire que les sentiments naissent sous le cortex est plausible au regard des données présentées plus haut : une destruction des cortex insulaires alors que les structures du tronc cérébral sont intactes est compatible avec un large éventail d'états sentimentaux ; les enfants hydrocéphales qui n'ont pas de cortex insulaire ni d'autres cortex somatosensoriels mais conservent des structures intactes dans le tronc cérébral ont des comportements témoignant d'états de sentiment.
Tout aussi important dans la génération des sentiments est un agencement physiologique central dans ma conception de l'esprit et du soi : le fait que les régions cérébrales impliquées dans la production de cartes corporelles et sous-jacentes aux sentiments forment une boucle de résonance avec la source même des signaux qu'elles cartographient. Dans le tronc cérébral supérieur, la machinerie chargée de la cartographie corporelle interagit directement avec la source même des cartes qu'elle forme, ce qui crée un lien étroit, une quasi-fusion entre le corps et le cerveau. Les sentiments émotionnels proviennent d'un système physiologique qui est sans parallèle dans l'organisme.
18 mars 2013 - Nos sentiments pourraient se passer de l’insula
Or voilà que le même Damasio vient de publier un article dans la revue Nature Neuroscience où il relativise la contribution de l’insula dans la genèse de nos sentiments. Il rappelle plusieurs observations qui ne sont pas très compatibles avec la thèse forte voulant que l’insula soit la plateforme essentielle de nos émotions et, par extension, de la conscience humaine qui s’élabore à partir de celles-ci.
Il rappelle d’abord que ce sont les nombreux noyaux de neurones de la partie supérieure du tronc cérébral qui reçoivent d’abord l’information en provenance du corps. Ces noyaux pouvant produire des représentations déjà élaborées de multiples paramètres de nos états corporels, leur contribution à nos sentiments ne peut pas être minimisée a priori.
Il pointe ensuite vers différents cas où l’insula est absente ou détruite, que ce soit les enfants qui naissent sans cortex cérébral ou des victimes d’encéphalites particulièrement dévastatrices. Or dans la plupart de ces cas, les gens conservent une riche palette d’états émotionnels.
D’où il conclut que la génération de nos sentiments doit forcément faire aussi appel au tronc cérébral et possiblement à nos cortex somatosensoriels SI et SII du lobe pariétal, des structures généralement épargnées chez les sujets à l’insula détruite qui conservent une riche vie émotionnelle.
Considérant d’autres sujets ayant subi des lésions corticales n’ayant pas affecté leur ressenti émotionnel, Damasio va même inclure une structure comme l'hypothalamus comme jouant probablement un rôle central dans la génération de nos sentiments, insistant ainsi sur les valeurs de survie évolutivement associées à la moindre émotion. Le substrat neuronal de nos sentiments les plus sophistiqués, de la compassion au dégoût en passant par le bonheur et la tristesse, pourrait s’enraciner dans des structures cérébrales présentes bien avant l’apparition des cortex cérébraux qui nous permettent aujourd’hui d’en faire d’exaltants ou de déchirants poèmes…
CORTEX FUSIFORME
Le gyrus fusiforme T4 (ou gyrus temporal latéral) est un gyrus de la face inférieure du lobe temporal du cortex cérébral. Ce quatrième gyrus temporal T4 est compris entre la scissure collatérale, en dedans, et la scissure temporo-occipitale, en dehors. Il se prolonge continument avec le lobule fusiforme O4 du lobe occipital pour former le gyrus temporo-occipital (ou gyrus occipito-temporal ou même gyrus fusiforme). Il est plus large dans sa partie moyenne.
(novembre 2009): Des experts ont démontré que le gyrus fusiforme n’était pas spécifique de la reconnaissance des visages (ou non reconnaissance : prosopagnosie), mais des reconnaissances expertes (dont les visages font parti); le gyrus fusiforme peut aussi être activé par les objets sur lesquels on a développé une expertise. Dans la condition «voiture» (où on présente de nombreuses voitures) : l'expert «voiture» a son gyrus fusiforme activé. Dans la condition oiseaux (où on présente de nombreux oiseaux): l'expert «oiseau» a son gyrus fusiforme activé.
CORTEX OCCIPITAL
Le lobe occipital se situe en arrière du cerveau (pôle postérieure). Il est divisé sur sa face interne par la scissure calcarine. Autour de la fissure calcarine se trouve l'écorce visuelle appelée également area striata ou zone 17 de Brodmann. Au niveau de l'écorce visuelle aboutissent les neurones provenant de la rétine (les fibres du nerf optique croisent partiellement la ligne médiane au niveau du chiasma optique). La moitié gauche du champ visuel se projette sur l'hémisphère droit et inversement. La destruction de l'écorce visuelle d'un côté entraîne une hémianopsie homonyme controlatérale. L'hémianopsie est la perte ou diminution de la vue dans une moitié du champ visuel d'un oeil. Le terme homonyme désigne une lésion ou un trouble concernant deux organes placés du même côté (tous deux à droite ou à gauche du plan médian). Fait important, la vision maculaire c'est-à-dire provenant d'une autre zone de la rétine est conservée. Ceci est dû au fait que la projection des neurones provenant de cette zone de la rétine se fait dans le cortex bilatéral (des deux côtés).
Les zones d'association visuelle correspondent aux zones 18 et 19 de Brodmann. Les lésions des zones d'association visuelle du côté de l'hémisphère qui domine (hémisphère gauche chez le droitier et inversement) entraîne des troubles de l'orientation dans l'espace et des troubles de la reconnaissance des objets dans une moitié du champ visuel, on parle dans ce cas d'agnosie visuelle.
CORTEX ORBITOFRONTAL ET VENTROMÉDIAN
Le cortex orbitofrontal est une région du cortex cérébral qui entre en jeu dans le processus de décision. Il est situé en position antérieure et sur la face inférieure du cortex préfrontal. Il prend son nom des lobes frontaux et du fait qu'il est situé au-dessus des orbites.
Cette partie du cortex préfrontal est en connexion avec le thalamus. Parce qu'il est actif dans les émotions et le système de récompense, le cortex orbitofrontal est souvent considéré comme faisant partie du système limbique.
le cortex orbito-frontal va associer les sensations émotionnelles, qui sont implicites et automatiques, à un stimulus. Il va en même temps enregistrer cette relation et sera, par ailleurs, capable de réactiver les sensations émotionnelles lors d'une rencontre ultérieure avec le stimulus conditionnant. Damasio (1995) parle de "marqueurs somatiques" qui permettraient donc à un individu de prendre en considération ses rencontres précédentes avec des stimuli et d'en tirer parti pour la réalisation de ses choix et de ses plans d'actions.
CORTEX PARIÉTAL
Le lobe pariétal est limité en avant par la scissure de Rolando et en bas par la scissure sylvienne. En arrière le lobe pariétal est limité par la scissure pariéto-occipitale. Cette zone du cerveau comprend l'écorce sensitive et la zone psycho sensorielle.
L'écorce sensitive c'est-à-dire la pariétale ascendante correspond aux zones 3, 1 et 2 de Brodmann. Il s'agit de la pariétale ascendante qui reçoit, par l'intermédiaire des neurones du thalamus, les sensations provenant des muscles, de la peau, des tendons et des articulations de l'organisme mais du côté opposé du corps. La destruction de la pariétale ascendante d'un côté s'accompagne dans la zone controlatérale, d'une anesthésie généralement non complète. L'irritation de l'écorce sensitive entraîne l'apparition de paresthésies (fourmillements) et de sensations douloureuses du côté opposé de l'organisme. On observe à la suite l'apparitions de crises d'épilepsie de type bravais-jacksonienne.
La zone psycho sensorielle se situe en arrière de la pariétale ascendante. À l'intérieur de la zone psycho sensorielle, qui est très complexe, s'élaborent les sensations qui sont également enregistrées à ce niveau. Les lésions de cette région entraînent une astéréognosie c'est-à-dire une perte de la possibilité pour un individu de reconnaître un objet en le touchant. On constate également une hémiasomatognosie (affection qui se caractérise par le fait que le patient refuse de reconnaître comme sien la moitié paralysée de son corps). Il s'agit du syndrome d'Anton-Babinski. Ceci est le résultat d'une atteinte pariéto temporale. L'enregistrement des mots écrits se fait à l'intérieur de la zone 39 et 40 de Brodmann. Une destruction de cette zone aboutit à l'impossibilité de lire et d'écrire. On parle dans ce cas d'agraphie et d'alexie.
CORTEX PRÉFRONTAL
A l’inverse des structures du système limbique qui dominent notre comportement lié aux émotions, le cortex préfrontal est en charge de notre capacité d’adaptation. C’est le cerveau de l’intelligence, de l’esprit d’initiative, de la prise de décision, du sang-froid. Des lésions de ces régions sont susceptibles d’induire des symptômes ressemblant à la dépression, comme la perte de flexibilité cognitive, le ralentissement psychomoteur et le manque de spontanéité, la dysphorie (inquiétude avec agitation maladive) ou l’anhédonie.
Les études d’imagerie cérébrale montrent une diminution de volume de la substance grise chez les sujets déprimés au sein de différentes régions cérébrales et en particulier le cortex préfrontal. De plus, on observe grâce à l’IRM fonctionnelle un hypofonctionnement au niveau préfrontal avec une diminution du flux sanguin et du métabolisme du glucose. Le cortex préfrontal n’est donc plus suffisamment actif pour contrecarrer nos réactions émotionnelles, il y a une rupture de l’équilibre avec l’amygdale hyperactivée. Les patients déprimés ne sont plus capables de faire face à leurs émotions, de prendre des décisions ou d’adapter leur comportement.
CORTEX TEMPORAL
Le lobe temporal repose dans la fosse moyenne du crâne. La scissure sylvienne sépare le lobe temporal du lobe frontal et du lobe pariétal. Le lobe temporal contient les aires auditives (écorce auditive : permettant d'entendre les sons) et plus précisément la zone 41 de Brodmann. Les hallucinations auditives qui surviennent quelquefois sont le résultat de lésions de type irritatif.
Le centre sensoriel du langage (centre de Wernick se situe dans l'os temporal à la partie postéro supérieur : gyrus supramarginalis). Les lésions survenant à ce niveau engendrent une impossibilité de comprendre le langage entendu alors que l'audition est normale. On parle dans ce cas d'aphasie sensorielle de Wernick.
L'écorce olfactive (sens de l'odorat) se situe également dans le lobe temporal, plus précisément au niveau de la partie interne du lobe temporal au niveau du gyrus uncinatus et de la circonvolution de l'hippocampe. Les hallucinations gustatives (quelquefois insupportables) apparaissent avant certaines crises d'épilepsie que l'on appelle les crises uncinées.
La moria est un trouble neurologique se caractérisant par une tendance à plaisanterie déplacée associée à une excitation et une tendance à la gaieté familière et bonhomme. Ceci survient au cours du syndrome du lobe frontal se caractérisant par une atteinte de la partie antérieure du lobe frontal qui comprend le tiers antérieur de l'hémisphère cérébrale.
Lobe temporal médian ou interne - LTM : (par JP Lachaux)
Nous en savons un peu plus sur le LTM depuis que le neurochirurgien Itzaak Fried et son équipe de chercheurs à Los Angeles ont entrepris d'enregistrer directement les neurones de cette région chez l'homme, lors des phases de repérage qui précèdent la chirurgie de l'épilepsie. Ces enregistrements leur ont permis de découvrir les neurones sensibles aux Beatles que j'ai déjà évoqués, et d'autres encore, fans de Jennifer Aniston, de Bal Clinton et de mère Teresa. Comme nous l'avons vu, ces neurones s'activent à l'évocation de concepts particuliers, quelle que soit la modalité utilisée, visuelle, auditive, verbale ou même imaginaire, puisque le simple fait pour le patient de penser aux Beatles ou à mère Teresa suffit à déclencher l'enthousiasme des neurones.
Dans une conférence récente, Fried nous a raconté l'histoire d'un neurone sensible à la moindre évocation des Simpsons, ce dessin animé américain très en vogue dans les années 1990. Le patient venait de regarder une série de petits extraits de films et de dessins animés, parmi lesquels figurait un extrait des Simpsons qui avait fait réagir ce neurone. Fried était curieux de savoir comment ce neurone réagirait à un simple souvenir ; et pour le savoir, il a demandé au patient de raconter ce qu'il avait vu : le patient a plongé dans ses souvenirs jusqu'à ce que le neurone se mette soudain à s'activer, une fraction de seconde avant que le patient ne déclare : « Ah oui... il y avait aussi un extrait des Simpsons. » Le neurone semblait avoir réagi à l'émergence du souvenir, comme si une petite bulle était remontée des profondeurs du cerveau. Il n'en faut pas beaucoup plus pour se convaincre que le LTM joue un rôle majeur dans l'imagination, et dans toutes ces promenades mentales qui occupent tant l'esprit pendant la douce rêverie et ce que nous appelons couramment la pensée.
Et ce n'est pas tout, car le rôle du LTM dans la construction et le rappel des souvenirs ne s'arrête pas là. En réfléchissant, il apparaît assez vite que le système de sélection attentionnelle, lorsqu'il choisit un objet, crée une dissociation entre deux types d'information : celles concernant cet objet, par exemple une tasse de thé posée devant moi, et celles concernant le contexte, l'environnement dans lequel il se trouve, une table, dans un café, etc. Quand mon attention se déplace vers la table, c'est au tour de celle-ci de devenir objet, tandis que la tasse se fond dans le contexte. Il y a donc toujours une séparation entre l'objet et son contexte. Au sein du cortex visuel, l'analyse de l'objet et celle de son contexte semblent impliquer deux systèmes différents, qui ne convergent qu'au sein du LTM, au sommet de la hiérarchie du système visuel. Selon des travaux récents, les informations relatives à l'objet parviendraient dans le LTM au niveau du cortex périrhinal et celles relatives au contexte au niveau du cortex parahippocampal. Le cortex entorhinal recevrait ensuite ces deux sources d'informations avant de les transmettre à l'hippocampe, qui associerait l'objet et son contexte pour former le souvenir.
Le rôle joué par le LTM dans la mémoire épisodique prend alors tout son sens, car chaque souvenir associe généralement un élément central et un contexte : je me souviens par exemple de l'ours polaire du zoo de Berlin, en janvier sous la neige. Je peux ensuite facilement imaginer ce même ours polaire sous la canicule du mois d'août, ou bien sur un iceberg en pleine mer, ou encore au sommet d'une montagne. En associant à chaque fois cette figure centrale avec un contexte différent, le nombre de scènes mentales que je peux créer est quasiment infini. Et ce n'est pas tout, car au sein du LTM, un système de connexions à double sens permet des allers-retours entre objet et contexte : un objet peut évoquer le contexte dans lequel il a été vu, et vice versa ; ainsi, le souvenir du zoo de Berlin me rappelle aussi celui des hippopotames, non loin de là, et celui de l'aquarium ; les souvenirs s'enchaînent pour reformer progressivement l'histoire de ce matin de janvier. Un lieu peut rappeler un événement qui y a été vécu ou un objet qui y a été vu. Le LTM a donc toutes les propriétés requises pour compléter le puzzle d'un souvenir à partir d'une seule de ses pièces.
Tout cela ne veut pas dire, bien sûr, que la pensée peut être ramenée à l'activité de quelques neurones dans le lobe temporal. Même dans le LTM, chaque concept n'active pas un neurone, mais, selon les estimations de l'équipe de Fried, des millions. De plus, chaque neurone réagit lui-même à l'évocation de nombreux concepts ; les observations de Fried montrent qu'un même neurone peut très bien réagir à l'évocation de la tour Eiffel et de la tour de Pise, et sans doute même à la vue d'une poutre en métal ou du drapeau français. Le LTM utilise ce que l'on appelle en neurosciences un codage neuronal par population; c'est-à-dire que chaque concept active une large population de neurones, dont chaque membre réagit également à d'autres concepts. La tour Eiffel et la tour de Pise activent donc deux populations de neurones qui coïncident partiellement.
Cette forme de codage permet à l'activité neuronale de se propager d'un concept à un autre comme un feu de forêt, selon le phénomène que nous appelons couramment associations d'idées. Si, parmi le million de neurones qui réagissent à la tour Eiffel, la moitié fait également partie de la population « tour de Pise », l'activité de ces cinq cent mille neurones peut se propager aux cinq cent mille restants et activer pleinement le concept « tour de Pise », pendant que l'activité des autres neurones « tour Eiffel » retombe tranquillement. Parmi le million de neurones « tour de Pise », six cent mille font peut-être partie d'une population « Italie », et ainsi de suite. Le sujet se perd dans ses pensées devant la tour Eiffel : « Elle me fait penser à la tour de Pise... l'Italie... Ce petit restaurant à Rome... Stella et sa robe rouge... Stella ! C'est l'anniversaire de Stella ! Il faut que je l'appelle ! » Cet homme n'est plus à Paris devant la tour Eiffel. Son attention vient d'être kidnappée par un processus neuronal tout à fait spontané et autoentretenu... C'est la troisième forme de captivation attentionnelle, la captivation cognitive. Bon voyage !
CORTICOÏDES ou CORTICOSTÉROÏDES
On donne le nom de corticostéroïdes (ou corticoïdes) aux hormones sécrétées par la région externe des glandes surrénales. Ils peuvent être séparés en trois groupes qui ont chacun leurs récepteurs distincts :
DÉPRESSION
La dépression entraîne au niveau cérébral des changements morphologiques et fonctionnels définissant la neuroplasticité ; ces changements s’opèrent au niveau du cortex préfrontal, de l'hippocampe et de l’amygdale et ont pu être révélés grâce aux progrès de la neuro-imagerie.
L’état dépressif est susceptible d’entraîner l’apparition de perturbations cognitives. Ces difficultés cognitives touchent essentiellement les capacités attentionnelles, la mémoire et les fonctions exécutives. Cette altération des fonctions cognitives est souvent associée à une intensité symptomatique plus élevée. Le déficit cognitif peut atteindre une intensité telle qu’il peut remplir les critères diagnostiques d’un état démentiel. Chez les patients souffrant de dépressions récurrentes, la plupart des études montrent que les déficits cognitifs sont plus sévères. Certaines des anomalies cognitives peuvent persister au-delà de l’épisode dépressif. En effet, plus de 2/3 des patients ont une altération résiduelle de la mémoire après traitement de l’épisode. Cela souligne l’importance d’évaluer les fonctions cognitives des patients après le retour à l’équilibre de l’humeur pour mieux caractériser l’évolution et la nature des symptômes non affectifs associés au trouble dépressif. Certains chercheurs font un lien direct entre les modifications structurales observées au niveau du cortex préfrontal et de l’hippocampe au cours d’un épisode dépressif et la détérioration des fonctions cognitives.
Les régions cérébrales qui se taisent ou s'emballent dans la dépression
Les connections entre le cortex ventromédian et le système limbique sont très denses. Elles en font donc une structure idéale pour lier le conscient à l’inconscient et pour donner un sens à nos perceptions en les liant en un tout significatif. De plus, cette région est modulée de façon importante par les neurotransmetteurs impliqués dans la dépression.
D’autres études portant sur des personnes ayant une forme familiale de dépression ou de maniaco-dépression ont montré que le cortex ventromédian pouvait être jusqu’à 40 % plus petit chez les personnes souffrant de dépression. En regardant au niveau cellulaire ce qui pouvait rendre compte de cette atrophie, on s’est rendu compte qu’au lieu d’observer une diminution de la quantité de neurones, c’était plutôt les cellules gliales qui étaient disparues en très grand nombre.
Or plusieurs expériences ont montré que ce « centre de contrôle des émotions » est presque inactif durant les phases de dépression (comme d’ailleurs plusieurs autres régions du cortex préfrontal) mais devient hyperactif durant les phases maniaques d’un trouble bipolaire. La logique de ces deux états opposés est aussi observable au niveau de certains symptômes comme celui, pour les personnes en phase maniaque, de trouver du sens dans tout ce qu'elles font, par opposition à la perte de sens globale qui afflige la personne en dépression.
Le cortex cingulaire est une autre structure cérébrale qui semble impliquée dans la dépression. Cette région ancienne du cortex est située sur la route de nombreuses voies remontant du cerveau émotionnel limbique inconscient ainsi que sur celle d’autres voies descendantes, conscientes, et impliquées dans la planification de l’action, en provenance du cortex préfontal.
Chez les personnes en dépression, on a observé une baisse d’activité dans le cortex cingulaire et, dans l’état inverse, la crise maniaque, on observe le contraire, soit une hyperactivité dans cette région.
La partie antérieur du cortex cingulaire, contrairement au reste de cette structure, devient quant à elle plus active durant une dépression. Bien que les fonctions du cortex (ou gyrus) cingulaire antérieur semblent très complexes, il semble qu’il agisse un peu comme une interface entre les régions inférieures plus instinctives du cerveau et les conduites plus rationnelles du néocortex. Son activation chez le sujet normal contribuerait ainsi à focaliser notre attention sur certaines choses. Des choses en provenance souvent de notre propre corps, comme la douleur par exemple ou les sentiments négatifs dans le cas de la dépression.
1) cortex orbitofrontal - 2) cortex préfrontal latéral - 3) cortex ventromédian - 4) système limbique - 5) cortex cingulaire antérieur
Une interprétation possible des études d’imagerie serait l’établissement d’un cercle vicieux entre diverses structures du système limbique suractivées durant la dépression.
une nouvelle voie thérapeutique ? (juillet 2012)
En France, 3 millions de personnes seraient touchées par la dépression. Cette maladie affecte de nombreux aspects de la vie quotidienne dont la joie de vivre, perdue chez de nombreuses personnes. La découverte de Rob Malenka, dont l’étude est publiée dans Nature, pourrait peut-être changer la vie des malades. L'équipe de ce neuroscientifique de l’Université de Stanford a mis en évidence le circuit de l’anhédonie, cette incapacité à ressentir des émotions positives lors de situations plaisantes. Il semblerait qu’il soit possible de jouer sur ce sentiment particulier. Cette découverte rend envisageable la mise au point de traitements spécifiques avec l’immense avantage dtés des autres symptômes liés à la dépression.
Le manque de plaisir identifié dans le cerveau
ENCODAGE
Pour le philosophe hollandais Baruch Spinoza, la mémoire acquiert de la force grâce à l'entendement et au raisonnement: « Plus une chose est intelligible, plus on la retient facilement et, au contraire, nous l'oublions d'autant plus facilement qu'elle l'est moins. Par exemple, si je donne à quelqu'un un grand nombre de mots sans lien, il les retiendra beaucoup plus difficilement que si je lui donne les mêmes mots sous la forme d'un récit." Cette conception inaugure des idées importantes connues sous le nom de « profondeur de l'encodage » (liée au traitement sémantique des éléments à mémoriser), qui donnera lieu à de nombreux travaux en psychologie cognitive et en neuropsychologie. Plus profond est le traitement lors de l'encodage de l'information et meilleure sera sa récupération ultérieure.
Les mécanismes d'encodage sont différents pour une information qui ne donne lieu qu'à un traitement superficiel et qui est oublié après quelques secondes, et pour une information qui est l'objet d'un traitement profond (sémantique), qui est intégrée aux connaissances préexistantes et conservée de façon durable.
Le modèle SPI (Seriel parallele indépendant) d'Endel Tulving prédit que l'encodage d'une information dans un système inférieur (la mémoire sémantique par exemple) peut se réaliser même si l'encodage dans le système supérieur (la mémoire épisodique) est défaillant, mais non l'inverse (ce qui est une contrainte forte).
L'encodage opère des transformations de l'information en provenance de stimulations externes ou résultant de traitements cognitifs et permet ainsi l'intégration des données sous un format de représentation mentale. La nature du traitement de l'information à l'encodage détermine la qualité de sa récupération. Selon la conception des niveaux de traitement proposée par Fergus Craik et Robert Lockhart en 1972, « la persistance de la trace mnésique est fonction de la profondeur de traitement, les niveaux de traitement profond étant associés à des traces mnésiques plus élaborées, plus résistantes dans le temps, et plus robustes ».
EPIPHYSE
Vous vous souvenez que Descartes pensait que la glande pinéale était le lieu où l'âme contrôlait le corps. Située à la base du cerveau, cette petite structure en forme de pomme de pin (d'où son nom) joue un rôle certes non négligeable, mais pas au sens métaphysique où Descartes l'entendait. La glande pinéale est responsable de l'analyse des informations concernant la clarté ou l'obscurité, nécessaires à l'ajustement de notre horloge biologique. Les fonctions cycliques comme le sommeil, l'éveil, les menstruations et le début de la puberté sont déclenchées par la glande pinéale. Cette glande produit également d'importantes substances, dont la sérotonine, l'histamine et la noradrénaline. De plus, c'est la seule source de mélatonine qui influe probablement sur l'horloge biologique.
EXOCYTOSE
Destinée d'une hormone dans la cellule neurosécrétrice.
FACTEUR NEUROTROPHIQUE
Les facteurs neurotrophiques sont des protéines très importantes pour le cerveau : sécrétées par les neurones eux-mêmes, leur territoire d'innervation, ou des cellules extraneuronales. Elles règlent la croissance, la prolifération et la différenciation des cellules nerveuses et sont ainsi essentielles au développement du système nerveux dans son ensemble, ainsi qu'au maintien des fonctions cérébrales. Ces propriétés ont rapidement intéressé les équipes de recherche travaillant sur la maladie de Parkinson, même si elle ne s'explique pas, à priori, par un déficit d'un facteur de croissance spécifique.
Trophique, trophicité : Ensemble des phénomènes qui conditionnent la nutrition et le développement d'un tissu (ensemble de cellules), d'une partie de l'organisme ou d'un organe.
FACTEUR DE CROISSANCE DU NERF (FCN)
Née en 1909, Rita Levi-Montalcini avait commencé ses études de médecine à Turin. Quand le gouvernement fasciste de Mussolini interdit aux juifs d'exercer la médecine et de faire de la recherche scientifique, elle s'enfuit à Bruxelles. Mais les Allemands menacèrent bientôt la Belgique, et elle revint à Turin où elle construisit un laboratoire secret dans sa chambre à coucher, afin d'étudier la formation des nerfs. Elle avait réussi à se forger des instruments de microchirurgie à partir d'aiguilles à coudre et commençait à obtenir des résultats quand les Alliés bombardèrent Turin. Elle dut fuir à nouveau, cette fois dans le Piémont. On était en 1940. Cette année-là, un jour qu'elle se rendait dans un petit village de montagne à bord d'un fourgon à bestiaux reconverti en wagon de voyageurs, elle tomba par hasard sur un article scientifique de Viktor Hamburger qu'elle dévora d'une seule traite et qui lui fit forte impression. Hamburger avait joué un rôle précurseur dans la compréhension des neurones en étudiant des embryons d'oeufs de poule. Elle décida de refaire ses expériences et se mit au travail sur une table dans un chalet de montagne, avec les oeufs d'un fermier voisin. Après chaque expérience, elle mangeait les oeufs. La guerre finie, Hamburger invita Levi-Montalcini à se joindre à son équipe de Saint Louis, qui avait découvert que la fibre nerveuse des poussins croît plus vite en présence de tumeur de souris. Levi-Montalcini soupçonnait cette tumeur de libérer une substance favorable à la croissance du nerf, et, avec le biochimiste Stanley Cohen, elle parvint à isoler la protéine responsable, qu'elle appela facteur de croissance du nerf, ou FCN. Le prix Nobel leur fut décerné à tous deux en 1986.
Les travaux de Rita Levi-Montalcini ont conduit à la découverte de plusieurs autres facteurs de croissance du nerf, parmi lesquels le facteur neurotrophique issu du cerveau, ou FNIC (En anglais BNDF (brain-derived neurotrophic factor). (N.d.T.)), qui retint toute l'attention de Merzenich.
FACTEUR NEUROTROPHIQUE ISSU DU CERVEAU (FNIC)
Le FNIC joue un rôle crucial dans la consolidation des changements plastiques qui affectent le cerveau pendant la période critique. D'après Michael Merzenich, il agit de quatre façons différentes.
Autisme et FNIC
FORMATION RÉTICULÉE
La formation réticulée (ou formation réticulaire ; également, en latin, formatio reticularis) est une structure nerveuse du tronc cérébral à l'interface des systèmes autonome, moteur et sensitif. Elle intervient dans la régulation de grandes fonctions vitales (comme les cycles veille-sommeil), le contrôle d'activités motrices réflexes ou stéréotypées, comme la marche ou le tonus postural et dans des fonctions cognitives telles que l'attention. De par son rôle central dans la régulation de la vigilance, les lésions de la formation réticulée entraînent souvent un coma.
La formation réticulée est située sur toute la longueur du tronc cérébral entre les grands faisceaux ascendants et descendants. Elle se présente sous la forme d'un tissu nerveux organisé dans son grand axe en trois bandes de corps cellulaires (médiane et para-médiane gauche et droite) eux-mêmes constitués en îlots de neurones densément connectés par de la substance blanche, d'où le terme réticulé qui vient du latin reticulum, réseau. Elle occupe les espaces laissés libres par les formations spécifiques (pédoncules cérébraux, protubérance annulaire, bulbe rachidien), mais n'a pas de limites anatomiques bien définies.
La partie de la formation réticulée que l'on appelle la formation réticulée activatrice ascendante (FRAA) reçoit des afférences sensitives et motrices et se projette à la fois sur le cortex cérébral et sur les noyaux du thalamus et de l'hypothalamus. La formation réticulée activatrice descendante projette des efférences vers le cervelet et vers les voies sensorielles. L’influence activatrice de la formation réticulée et autres structures sous-corticales sont transmises au cortex cérébral par deux types de projections : la voie thalamocorticale et la voie extrathalamique.
Phylogénétiquement, la formation réticulée est l'une des plus anciennes structures nerveuses du tronc cérébral, lui-même étant ce qu'on peut considérer comme un des centres les plus primitifs du système nerveux central.
Elle joue un rôle important dans la douleur.
Noyaux de la formation réticulée paramédiane
Le tiers latéral est une structure associative qui est atteinte par des autres fibres du tronc (région latérale ) et des fibres cortico-réticulaires (cortex prémoteur).
La partie médiane est la localisation des fibres ascendantes et descendantes. Ces noyaux, grâce à leurs nombreuses projections ascendantes et descendantes, sont impliqués dans le " système réticulé activateur ascendant " qui projettent l'ensemble du système nerveux central.
Ces noyaux, grâce à leurs nombreuses projections ascendantes et descendantes, sont impliqués dans le " système réticulé activateur ascendant " qui projettent l'ensemble du système nerveux central.
Ces noyaux sont représentés (de la région caudale à la région crâniale) par :
Noyaux de la formation réticulée latérale
Les noyaux de cette région sont constitués de cellules de petite taille qui sont implqués dans de nombreux réflexes des nerfs crâniens et dans les régulations viscérales comme la respiration ou la circulation…
Ces noyaux sont représentés (de la région caudale à la région crâniale) par :
Dans cette partie latérale, on trouve également des noyaux (de la région caudale à la région crâniale) dont les fibres se projettent sur le cervelet :
HIPPOCAMPE
L'information en provenance des cortex associatifs visuels, auditifs ou somatiques parvient d'abord à la région parahippocampique du cortex, puis passe au cortex enthorinal et finalement à l'hippocampe comme telle. À travers celui-ci, l'information va passer successivement à travers trois régions distinctes.
L'hippocampe, les structures corticales qui l'entourent, ainsi que les voies nerveuses qui les relient à l'ensemble du cortex sont grandement impliquées dans la mémoire déclarative, celle des faits et des événements.
L’hippocampe est la structure cérébrale dont les liens avec la symptomatologie dépressive ont été les mieux démontrés. L’hippocampe participe à des fonctions aussi essentielles à la vie relationnelle que la régulation de l’humeur, l’acquisition des connaissances et de façon plus générale, à l’adaptation d’un individu à son environnement. Chez l’homme, des dommages hippocampiques bilatéraux entraînent une amnésie antérograde, ce qui suggère l’implication de l’hippocampe dans l’acquisition de nouvelles informations. Il a une fonction spécifique à la mémoire déclarative, dans laquelle un effort conscient de souvenir est requis.
(voir aussi : stress)
Les études d’imagerie morphologiques, montrent une diminution du volume de l’hippocampe chez les sujets déprimés et ce, indépendamment de l’âge et du sexe. De plus, la fréquence des épisodes dépressifs et la durée de l’épisode passée sans traitement antidépresseur sont inversement corrélées à la taille de l’hippocampe : plus la durée des troubles est longue, plus l’hippocampe est petit. On retrouve d’ailleurs des troubles de la mémoire narrative chez les patients déprimés, qui sont eux-mêmes corrélés à la durée totale passée en dépression objectivant de manière clinique ces altérations anatomiques.
Le gyrus denté
Définition : Le gyrus denté ou gyrus dentatus (ou gyrus dentelé, corps godronné) est un gyrus du lobe limbique du cortex cérébral. Il est situé au dessus du sillon hippocampique, le long de l'hippocampe. C'est un cylindre de substance nerveuse plicaturé (formant de petites dents) qui se prolonge en avant, par la bandelette ou limbus de Giacomini enjambant l'uncus et à l'arrière, par le faisceau gris (fasciola cinerea).
HOMÉOSTASIE
Le concept d'homéostasie explique la constance du milieu intérieur et sert de fondement théorique à la physiologie des régulations. L'ensemble des grandeurs régulées définit la constance du milieu intérieur. Les plus importantes sont le contenu en gaz du sang, l'acidité ou PH, la température, le taux de sucre, la pression artérielle, la pression osmotique.
(C’est la force qui attire l’eau au travers de la membrane qui sépare deux milieux liquides. Le créateur de cette force c’est le sel qui attire l’eau.) Plus une solution est salée, plus sa pression osmotique est élevée. Si, pour quelque raison, l'animal perd de l'eau la concentration de sel dans le milieu intérieur, autrement dit la pression osmotique s'élève.
Les constantes obéissent à une hiérarchie. Les plus importantes doivent être maintenues à tout prix, parfois en sacrifiant une constante subalterne.
HORMONES voir aussi : exocytose
LE CERVEAU HORMONAL
Les neurones du cerveau hormonal se distinguent de ceux du cerveau câblé de plusieurs façons. Regroupés surtout dans le tronc cérébral et la région centrale du cerveau, ils forment de petits amas de milliers de cellules qui projettent toutefois leurs axones dans de vastes régions de l'encéphale.
Un seul de ces neurones peut donc en influencer plus de 100 000 autres grâce aux neuromodulateurs qu'ils déversent dans l'espace extracellulaire plutôt que dans la fente synaptique.
Il en résulte aussi des effets plus long à s'établir et qui durent plus longtemps que ceux des neurotransmetteurs des circuits du cerveau câblé. L'implication de seconds messagers dans le mode d'action des neuromodulateurs n'est pas étranger à cette dynamique.
Les quatre neuromodulateurs principaux sont : la noradrénaline, la sérotonine, l'acétylcholine et la dopamine.
Deux groupes de faits expérimentaux montrent ce cerveau hormonal en action. L'injection d'une quantité infime d'une hormone peptidique dans une région déterminée du cerveau déclenche un comportement intégral avec toutes ses séquences motrices et adaptatives. C'est le cas pour l'angiotensine, qui provoque un comportement de boisson, de l'ocytocine dont l'injection dans l'hypothalamus d'une ratte est suivie de l'apparition chez celle-ci d'un comportement maternel caractérisé et, enfin, de la lulibérine, qui induit un comportement sexuel (voir plus bas). Ces expériences ont été effectuées sur des rats, mais certains auront vite fait la transposition à l'homme. Je m'empresse de dire aux amateurs du courrier du cœur, qui doivent être chagrinés par un réductionnisme aussi trivial, que je ne vois pas pourquoi la plus exquise chimie des sentiments ne pourrait pas coexister dans le cerveau avec l'affinité d'une hormone pour ses récepteurs.
Il est important de noter que la même hormone, qui provoque dans le système nerveux le comportement, intervient à la périphérie dans la régulation homéostasique correspondante : angiotensine et volume sanguin, lulibérine et ovulation, ocytocine et lactation. On sait, par ailleurs, que l'hormone cérébrale et l'hormone périphérique proviennent de sources différentes : dans le milieu cérébral, le cerveau sécrète de l'angiotensine, de la lulibérine et de l'ocytocine, au même titre que le rein et l'hypophyse secrètent les hormones périphériques. La neuro-humeur cérébrale et l'hormone périphérique participent donc à la même homéostasie et tout se passe comme si une r@?gulation nerveuse centrale utilisant les mêmes agents chimiques venait, en quelque sorte, doubler les régulations hormonales systémiques avec la finesse adaptative que lui confère la complexité cérébrale. Le corps et son double, le cerveau.
Un deuxième groupe de faits expérimentaux récents nous ramène aux concepts de base de l'endocrinologie. Ce sont les greffes de fragments de cerveau au sein de territoires cérébraux déficients. La réussite fonctionnelle de ces greffes capables, dans certains cas favorables, de restaurer des fonctions cognitives (apprentissage, exploration de l'espace etc) est proprement sidérante en regard de la conception d'un cerveau câblé, qui tirerait son sens de la précision de ses connexions. La greffe n'est que la suite logique de la démarche thérapeutique, qui consiste à soigner des malades atteints de troubles moteurs (maladie de Parkinson) par l'administration du neurotransmetteur manquant (DOPA) ; elle ne diffère pas fondamentalement de l'opothérapie* des premiers endocrinologues.
Le cerveau peut être considéré comme une glande dans toute l'acception du terme. A côté de la sortie nerveuse par les neurones moteurs qui commandent muscles et viscères, il existe une sortie hormonale au niveau d'un entonnoir formé par le plancher du cerveau, l'hypothalamus. Les produits de sécrétion de l'hypothalamus obéissent aux deux principes fondamentaux qui définissent une hormone : action à distance et autorégulation par rétroaction. Certaines hormones sont destinées à agir sur des organes périphériques comme le rein ; d'autres agissent très localement sur l'hypophyse dont elles contrôlent les sécrétions. Par l'intermédiaire de ces hormones hypophysiotropes, les sécrétions hypothalamiques s'adressent à l'ensemble des glandes de l'organisme. La cellule hypophysaire apparaît comme un véritable noeud stratégique dans l'interdépendance cerveau-organisme. Ces hormones du cerveau exercent leur action inhibitrice ou stimulatrice sur les différents types de cellules hypophysaires. Il s'agit d'une régulation multifactorielle, chaque hormone hypophysaire étant sous la dépendance de plusieurs agents d'origines cérébrale et périphérique et un même agent intervenant dans la régulation de plusieurs hormones. Cette multiplicité permet un spectre étendu de réponses neuroendocrines qui s'intègrent dans l'ensemble des régulations adaptatives. Enfin, il est par ailleurs remarquable que la cellule hypophysaire rassemble les caractéristiques d'une cellule endocrine et d'une cellule nerveuse. A titre de revanche, les méthodes de l'électro-neurophysiologie sont aujourd'hui applicables à la cellule hypophysaire qui s'est révélée excitable et capable de générer des influx électriques.
Chimie des passions
1 - La lulibérine :
Prenez un individu normalement constitué et, grâce à une fine canule introduite dans son cerveau, saupoudrez une étroite parcelle de son hypothalamus d'une pincée de lulibérine : votre cobaye, pour peu qu'il ait une partenaire bien disposée à sa portée, se livrera sur l'heure à une violente et répétitive activité amoureuse. L'histoire est inquiétante, mais soyez rassurés, l'individu en question n'est qu'un rat de laboratoire, et il est peu probable qu'Iseult ait usé de lulibérine pour rendre Tristan amoureux : la barque du roi Marc n'était pas un laboratoire de psychophysiologie.
Il n'empêche, le fait expérimental est là. Il suffit de l'injection, dans une région déterminée de l'encéphale, d'une infinitésimale quantité d'un certain peptide pour déclencher chez l'animal la séquence complète de son comportement amoureux depuis les travaux d'approche jusqu'à la consommation de l'acte.
Mais l'aventure chimique ne s'arrête pas là. Des dosages ont montré que l'explosion finale du coït s'accompagnait d'une libération massive d'endorphines. Ces peptides seraient responsables, par leur action inhibitrice sur les cellules nerveuses de l'hypothalamus, de la satiété sexuelle.
Alors, le désir, l'amour, la faim : la libération en cascade de substances chimiques à l'intérieur du cerveau ? Nous ne serons pas assez naïfs pour succomber à la tentation d'un réductionnisme si peu romantique. Certains discours mythologiques clamés sur les rivages psychanalytiques ne se privent d'ailleurs pas d'identiques simplifications. Que savons-nous, après tout, des peptides sécrétés par le cerveau du jeune Œdipe lorsqu'il fut privé du sein de sa mère ? Ce qui nous intéresse, c'est de savoir qu'une substance unique appliquée expérimentalement ou libérée naturellement dans le cerveau est capable de déclencher la séquence complète et ininterrompue des actes moteurs infiniment complexes et variés qui constituent un comportement déterminé.
2 - La vasopressine :
si, pour quelque raison, l'animal perd de l'eau, la concentration de sel dans le milieu intérieur, autrement dit la pression osmotique, s'élève. Variable régulée, la pression osmotique sera maintenue constante par la mise en jeu de mécanismes régulateurs : freiner les sorties d'eau ou/et augmenter les entrées. Diminuer les sorties, c'est freiner l'élimination rénale : une hormone sécrétée par le cerveau, l'hormone vasopressine, encore appelée hormone antidiurétique, se charge de la besogne. Le taux de l'hormone antidiurétique circulant dans le sang est une variable contrôlée qui augmente en réponse à l'élévation de la pression osmotique : exemple de régulation hormonale. Augmenter les entrées, c'est boire : l'élévation de la pression osmotique du sang au-delà d'un seuil déclenche immédiatement une sensation de soif et le besoin impérieux de boire, type même de régulation comportementale. Cet exemple montre la diversité des mécanismes régulateurs qui peuvent être de deux sortes : hormonaux et comportementaux. Un chameau, malgré la sécheresse du désert, n'a pas une pression osmotique beaucoup plus élevée que celle d'un patron de bistrot, il dispose seulement de mécanismes régulateurs plus puissants et adaptés au milieu extérieur qui autorisent des écarts considérables des variables contrôlées : diurèse et boisson.
3 - L'angiotensine :
On connaît cette soif inextinguible des blessés qui saignent. On sait aujourd'hui qu'elle est due à la libération d'une hormone peptidique appelée angiotensine. Celle-ci est fabriquée en collaboration par le foie et le rein pour lutter contre une diminution brutale du volume sanguin, lors d'une hémorragie par exemple. Cette hormone a pour effet de contracter les vaisseaux et d'adapter le contenant vasculaire au contenu sanguin diminué, évitant ainsi une chute préjudiciable de la pression artérielle. L'injection de quelques milliardièmes de gramme d'angiotensine dans des régions précises du cerveau déclenche instantanément chez l'animal un comportement de boisson. Même chez un animal gavé d'eau jusqu'à plus soif, la boisson devient la source exclusive de son désir. Comme avec la lulibérine, la séquence déclenchée est complète et se déroule de façon inéluctable depuis la quête irrépressible d'eau jusqu'à sa prise compulsive et prolongée.
4 - L'ocytocine :
Est-il spectacle plus attendrissant qu'une mère soignant ses petits ? A sa vue, le coeur du spectateur le plus endurci plonge dans un sentiment de communion avec la nature, la bête et le sublime réconciliés !
L'injection d' ocytocine — hormone hypophysaire qui fait jaillir le lait des mamelons — dans les ventricules cérébraux d'une rate vierge provoque en quelques minutes un comportement maternel auquel ne manque aucun composant : bricolage hâtif d'un nid, regroupement et recouvrement de ratons étrangers placés dans la cage, léchage et récupération des petits égarés.
HYPOPHYSE
L'hypophyse, au centre du cerveau, a la taille d'un haricot. Elle est accolée à l'hypothalamus et produit nombre d'hormones importantes comme l'ACTH (adrenocorticotropic hormone en anglais), qui joue un rôle dans les réactions d'alerte.
Par le passé, l'hypophyse était considérée comme la glande la plus importante du corps humain, mais nous savons maintenant qu'elle opère sous l'influence de l'hypothalamus, qui la relie au cerveau. L'hypothalamus répond aux besoins organiques, soit par des influx nerveux, soit par induction de l'hypophyse. Celle-ci produit alors des hormones qui circulent vers les organes via le flux sanguin.
Comme tant d'autres parties du cerveau, l'hypophyse présente deux lobes : antérieur et postérieur. Chacun libère des hormones différentes qui jouent un rôle sur la croissance et régulent l'activité d'autres glandes.
L'hormone de croissance produite par l'hypophyse influe sur la taille. Si cette hormone est en surproduction, cela peut provoquer un gigantisme chez l'enfant et une acromégalie (du grec ancien akros, haut, extrême, et megalos, grand de taille) chez l'adulte. L'acromégalie est une maladie qui se caractérise par une croissance exagérée du visage et des extrémités. À l'opposé, si cette hormone est en sous-production, cela provoque le nanisme.
Quelle est la différence entre un joueur de basket de 2,20 m et une personne souffrant d'acromégalie ? Le joueur de basket a un taux d'hormones de croissance légèrement supérieur à la moyenne, alors que le patient atteint d'acromégalie présente un taux d'hormone en augmentation anormale et persistante. Ce phénomène est généralement dû à une tumeur bénigne nommée « adénome pituitaire ».
HYPOTHALAMUS
L'hypothalamus, structure qui pèse 5 grammes et a la taille de deux petits pois, est le thermostat du corps. Il contrôle la plupart des fonctions vitales, comme la respiration, le battement cardiaque, la digestion, l'alimentation, l'équilibre hydrique, la température corporelle, la sécrétion d'acide gastrique ou le sommeil, ainsi que certaines fonctions endocrines (qui sécrètent des hormones). Vous pourriez vous dire : « D'accord, ces fonctions sont importantes, mais ce n'est pas ce qu'il y a de plus excitant dans l'existence. » Sauf que la petite boule de nerfs sous le thalamus est également impliquée dans l'émotion et l'activité sexuelle.
L'hypothalamus comporte un dipsostat (de dipso, « je bois ») localisé dans les noyaux supra-optiques. Lorsque la pression osmotique des liquides de l'organisme augmente (augmentation de la salinité du milieu intérieur), lorsque le sang est hémoconcentré par perte de plasma, la sensation de soif apparaît. L'hypothalamus capte le phénomène grâce à des osmo-récepteurs, et en réponse, d'une part réduit la diurèse en libérant par l'intermédiaire de l'hypophyse postérieure l'hormone antidiurétique (ADH) qui agit sur le rein, d'autre part induit chez l'animal une conduite de recherche de l'eau. Lorsqu'on stimule électriquement les noyaux supra-optiques d'une chèvre, on provoque chez elle une véritable pulsion à boire avec ingestion d'eau qui peut aller jusqu'à 40 % du poids de l'animal, pendant tout le temps que dure la stimulation.
L'hypothalamus comporte un orexostat (d'orexo, je mange) localisé dans les noyaux ventro-médians et dans l'hypothalamus latéral.
Les noyaux ventro-médians de l'hypothalamus sont des centres de la satiété. Ils reçoivent directement leurs impressions du tube digestif. Les signaux issus de l'estomac rempli réduisent vite la pulsion de faim. La destruction expérimentale de ces noyaux chez l'animal entraîne une boulimie bientôt suivie d'une obésité qui peut devenir monstrueuse.
Les noyaux de l'hypothalamus latéral sont des centres de l'appétit. Leur destruction chez l'animal conduit celui-ci à l'anorexie totale. La mort survient sauf si l'alimentation est maintenue par sonde gastrique. Au contraire leur stimulation provoque une conduite forcenée de recherche de la nourriture, une véritable agression à la recherche d'une proie à dévorer. Le niveau du sucre sanguin (glycémie) est certainement important dans le déclenchement ou l'arrêt de telles conduites (d'où les fringales du diabétique en hypoglycémie parce qu'il a un peu trop forcé sur sa dose quotidienne d'insuline, et peut-être l'expression populaire « avoir les crocs » qui peut rendre compte de la mauvaise humeur, des colères, de l'agressivité du sujet à jeun).
Chez l'homme, et même chez l'animal, les faits sont en réalité moins simples : il n'y a pas exactement de centre de la satiété, ni de centre de l'appétit. Des circuits neuronaux plus complexes font intervenir l'odorat, le goût et leur projection corticale. Privé d'odorat, un rat consomme peu à la fois mais sans interruption (« consommation émiettée continue »). L'hypothalamus règle aussi, il faut s'en souvenir, le niveau des réserves de graisses (lipostat ?). Alors que le boa qui vient d'avaler un chevreau reste spontanément trois semaines sans déjeuner, l'homme fait des « hyper-repas ». Sa conduite alimentaire est stimulée par l'agréable ou inhibée par le désagréable. Ainsi, la forte « palatabilité » du « régime cafétéria » met en défaut ses régulations hypothalamiques. La correction métabolique n'intervient plus guère et manger devient pour lui « activité esthétique » (Le Magnen). Il a perdu ce que Cannon nommait joliment « la sagesse du corps ».
L'hypothalamus est un gonadostat (gonades : glandes sexuelles) localisé dans les noyaux postérieurs, centres de contrôle et de régulation de la sexualité et de la reproduction qui créent les pulsions correspondantes et guident la conduite de recherche d'un ou d'une partenaire. L'hypothalamus est le relais indispensable à la réalisation de toute conduite sexuelle, de la parade nuptiale à la copulation et à l'éjaculation chez le mâle (acte consommatoire au même titre que la satisfaction de la soif ou de la faim). L'assouvissement de ces besoins fondamentaux assure la survie de l'individu et de l'espèce. Des conduites de maternage, l'allaitement chez les mammifères sont programmés à ce niveau. C'est cette m@?me région de l'hypothalamus postérieur qui tient sous sa dépendance, par la sécrétion de neuro-hormones excitatrices, le fonctionnement de l'hypophyse (aire dite « hypophysiotrope »). Un rétro-contrôle sensible au taux sanguin des hormones hypophysaires circulantes joue ensuite pour réduire ou stopper l'activité de cette aire de l'hypothalamus.
Ainsi cette extraordinaire structure qu'est l'hypothalamus préside à presque toutes les régulations endocriniennes.
Les noyaux hypothalamiques
IMAGERIE BIOMÉDICALE : IRM TEP et EEG
La troisième invention majeure de l'imagerie médicale (après le CT-scan ou CAT-scan et le PET-scan) est l'imagerie par résonance magnétique ou IRM, qui a été utilisée pour la première fois sur des humains en1977. Ses inventeurs sont Raymond Damadian, Larry Minkoff et Michael Goldsmith.
La première machine IRM a été appelée l'Indomptable, car elle était l'aboutissement de sept ans d'efforts et beaucoup la jugeaient irréalisable. Le prototype de la machine est conservé à la Smithsonian Institution à Washington.
L'IRM est un puissant instrument diagnostique. Les détails sont encore plus grands que dans les autres techniques d'imagerie et le médecin peut adapter l'engin à un problème spécifique. L'IRM utilise un aimant géant en forme de tunnel, dans lequel est placé le patient (le champ magnétique crée par cet aimant est 30000 fois plus puissant que celui de la Terre, voire aujourd'hui 90 000 fois). Des ondes électromagnétiques provoquent une aimantation des protons dans le corps et les informations recueillies sont enregistrées dans l'ordinateur, ce qui permet la visualisation d'images selon des angles et des directions variés.
L'examen IRM est indolore, mais il peut être désagréable et même provoquer l'anxiété chez les sujets claustrophobes. Le patient qui passe une IRM est allongé sur une table qui coulisse à l'intérieur d'une sorte de cercueil cylindrique. La procédure dure en général entre 20 et 30 minutes et le bruit de la machine est si énorme que le patient doit mettre des boules Quiès, qui atténuent mais n'éliminent pas complètement le martèlement. La bonne nouvelle, c'est que les médecins prescrivent habituellement un léger calmant pour apaiser le patient.
Autre défaut de l'IRM, c'est une technique très coûteuse et qui n'est donc pas disponible dans tous les hôpitaux.
IRM : appareil capable de détecter le champ magnétique des protons (vous savez les constituants de l'atome, proton, électron, neutron). En effet, les protons des atomes de la matière vivante sont animés d'un mouvement de rotation (spin) qui crée un champ magnétique. Placée dans un gros aimant (le tunnel de l'hôpital), la perturbation magnétique peut être enregistrée et donner une image. En neuropsychologie, l'IRM est utilisée pour suivre les globules rouges du fait d'un afflux sanguin dans les zones du cerveau qui sont actives. Ce n'est autre que l'hémoglobine, la protéine transportée par les globules rouges qui est chargée de convoyer l'oxygène depuis les poumons jusqu'aux tissus de l'organisme. Plus précisément, l'hémoglobine existe sous deux formes : elle arrive dans les tissus chargée d'oxygène (c'est l'oxy-hémoglobine), elle leur en abandonne une partie et se transforme alors en déoxy-hémoglobine. Or l'oxy-hémoglobine est invisible en IRM alors que la déoxy-hémoglobine a pour effet de noircir l'image. Quand un petit volume de cerveau augmente son activité la quantité de sang augmente avec majoritairement de l'oxy-hémoglobine d'ou une diminution globale de la déoxy-hémoglobine et donc une augmentation du signal IRM. En bref, cela se traduit par une teinte localement plus claire de l'image.
Ainsi peut-on voir en « temps réel » l'activation des zones du cerveau impliquées dans les tâches cognitives en jaune ou rouge), ou au contraire les zones lésées (en bleu ou violet) (voir Houdé et al., 2002 pour de nombreux exemples, ou sur Internet, par exemple sur le site du CEA).
Grâce à l'IRM, il est maintenant possible de voir non seulement la forme du cerveau, mais aussi son activité, sans jamais avoir à ouvrir le crâne – on parle alors d'IRM fonctionnelle, ou d'IRMf. L'IRMf voit le cerveau comme un assemblage de milliers de petites briques de Lego de quelques millimètres de côté, appelées voxels, dont elle sait mesurer la consommation d'oxygène. Comme les neurones ont besoin d'oxygène pour travailler, l'IR/vIf permet d'identifier avec précision les régions du cerveau dont l'activité augmente, ou diminue, quand une personne est occupée à faire quelque chose – qu'il s'agisse de lire attentivement un livre ou de s'imaginer en train de jouer au tennis. L'IRMf fournit donc des cartes d'activité cérébrale en trois dimensions.
En comparant pour chaque voxel la consommation d'oxygène du cerveau pendant une tâche cognitive et pendant sa condition contrôle, il est maintenant possible de localiser au millimètre près les zones cérébrales associées à un processus cognitif.
Quand à la TEP (tomographie par émission de positons), on se croirait projeté dans un film de science-fiction, car il s'agit... d' antimatière !!! En cassant des atomes dans les accélérateurs de particules (ex. le CERN à Genève), les physiciens avaient découvert que certaines particules atomiques disparaissaient pour donner place à des rayonnements (photons). Pour expliquer ce mystère, ils ont imaginé que la particule rencontrait son antiparticule et était ainsi annihilée en donnant de l'énergie ; ainsi naissaient les antiprotons, les positons (antimatière de l'électron). Eh bien la TEP est basée sur ce principe et utilise les positons. TEP signifie tomographie (c'est-à-dire « couper en tranches » du grec tomo) par émission de positons : un atome radioactif est injecté par voie veineuse (on utilise souvent de l'eau contenant un atome d'oxygène radioactif : O15 au lieu de l'atome d'oxygène ordinaire.) et en se désintégrant, il émet des positons qui parcourent quelques millimètres avant de rencontrer des électrons pour s'annihiler en produisant des photons (rayons gamma) qui sont photographiés par tranches fines, ce qui donne une image tridimensionnelle des organes.
Électroencéphalographie - EEG
voir aussi La magnétoencéphalographie (MEG)
INTERACTION CERVEAU/CORPS
A.D. (L'erreur de Descartes) : ..il existe des interactions bien connues entre le cerveau et le corps, et on peut aussi mentionner des interactions, peut-être moins visibles, entre le fonctionnement mental et le corps. Soit l'exemple suivant :
Bien entendu, on a également observé l'inverse, c'est-à-dire l'influence sur le cerveau de substances chimiques provenant du corps. Nul ne sera surpris d'apprendre que la nicotine, l'alcool et les substances pharmacologiques (médicales ou non) peuvent pénétrer dans le cerveau et modifier son fonctionnement, et ainsi altérer les phénomènes mentaux. L'action des substances chimiques en question peut concerner directement les neurones ou leurs cellules de soutien. Ou bien elle peut être indirecte et dépendre des neurotransmetteurs libérés par les neurones modulateurs localisés dans le tronc cérébral et la base du télencéphale, que nous avons envisagés dans le chapitre préc@?dent. Lorsqu'ils sont activés, ces petits ensembles de neurones peuvent déverser une certaine quantité de dopamine, de noradrénaline, de sérotonine ou d'acétylcholine dans de vastes régions du cerveau, comprenant notamment le cortex cérébral et les ganglions de la base.
LIQUIDE CÉPHALO-RACHIDIEN OU CÉRÉBRO-SPINAL
Dans un cerveau de 1 400 grammes, les cavités emplies de liquide représentent à peu près 100 millilitres, soit la valeur de deux verres à bordeaux (figure 12). Au coeur de chaque hémisphère, les ventricules latéraux contiennent la plus grande partie de ce liquide céphalo-rachidien. Ils communiquent par un orifice étroit avec le troisième ventricule, pièce d'eau médiane en forme d'entonnoir autour de laquelle s'ordonnent les structures nerveuses où se jouent les passions : cerveau limbique et hypothalamus. Cet espace communique vers l'arrière par l'aqueduc de Sylvius avec le quatrième ventricule, bassin losangique dont le plancher recouvre les centres vitaux — respiratoires, cardiaques et vasculaires — et dont le toit forme la base du cervelet. Un canal étroit prolonge ce dernier ventricule à l'intérieur de la moelle épinière et, par des orifices situés à son extrémité inférieure, le fait communiquer avec les espaces qui entourent le cerveau et la moelle ; ces orifices permettent au liquide de se répandre sur la convexité des hémisphères, dans la profondeur des sillons, tout en baignant la base du cerveau et la moelle sur toute sa hauteur. Les fontaines qui alimentent les espaces liquidiens sont placées à l'intérieur des ventricules latéraux. Ces plexus choroïdes sont formés de vaisseaux capillaires recouverts de cellules épithéliales jointives qui tirent du plasma sanguin et sécrètent dans les cavités céphaliques un liquide dont la composition est celle du milieu extracellulaire cérébral. Avec un débit de 0,35 millilitre par minute, les plexus choroïdes font sourdre environ un demi-litre de liquide céphalo-rachidien à l'intérieur des ventricules latéraux, d'où il se répand, par l'intermédiaire des ventricules médians, vers la base et la convexité du cerveau.
Le liquide des cavités se répand dans celui qui baigne les cellules nerveuses. Aucune barrière ne s'oppose à la diffusion des substances du liquide céphalo-rachidien dans l'espace extracellulaire cérébral. En retour, les produits de sécrétion des neurones ont la possibilité de se répandre grâce au liquide céphalo-rachidien dans l'ensemble des structures cérébrales. Ce liquide peut être considéré comme un véritable système d'alimentation et de drainage du cerveau. Il offre un support à ce que Rémy Colin a appelé l'hydrocrinie3, c'est-à-dire à la diffusion d'hormones par voie liquidienne au sein du milieu cérébral. Le liquide sécrété est repris par la circulation veineuse au niveau de formations granulaires, sortes de petites valves permettant l'échappement dans le sang du liquide céphalorachidien et des solutés. Il est important d'insister sur l'étanchéité des plexus choroïdes et des valvules de sortie qui n'interrompent donc pas la barrière hémato-encéphalique, alors qu'en revanche il existe un libre passage du liquide céphalo-rachidien vers l'espace extracellulaire cérébral.
LOCALISATIONNISME
Si le cerveau peut se réorganiser, il s'ensuit que le localisationnisme ne peut en offrir une image appropriée. Au début, même Paul Bach-y-Rita était localisationniste, à cause des progrès accomplis par cette théorie. Le localisationnisme scientifique est né en 1861, quand le chirurgien français Paul Broca se trouva confronté à un patient victime d'une attaque cérébrale qui avait perdu la faculté de parler et ne pouvait prononcer qu'un seul mot. Quoi qu'on lui demandât, le malheureux répondait : « Tan, tan. » À sa mort, Broca disséqua son cerveau et trouva des tissus lésés dans le lobe frontal gauche. Les sceptiques doutaient qu'on puisse localiser la fonction langagière dans une zone unique jusqu'à ce que Broca leur montre les tissus endommagés et fasse le lien avec d'autres malades frappés d'aphasie et pr@?sentant des lésions dans la même zone cérébrale. Cette zone fut bientôt appelée « zone de Broca », et l'on supposa qu'elle avait pour fonction de coordonner les mouvements musculaires des lèvres et de la langue. Peu après, un autre médecin, l'Allemand Carl Wernicke, corréla des lésions d'une autre partie du cerveau à un problème distinct : l'incapacité à comprendre le langage. Wernicke émettait l'hypothèse que la zone endommagée était en charge des représentations mentales des mots et de leur compréhension. On l'appela « zone de Wernicke ». Par la suite et durant près d'un siècle, le localisationnisme s'ancra dans les esprits au fur et à mesure que de nouvelles recherches perfectionnaient la cartographie du cerveau.
Mais malheureusement, on en vint assez vite à extrapoler, et le localisationnisme, d'une série de corrélations intrigantes basées sur les rapports entre des lésions circonscrites à une région cérébrale et la perte de fonctions mentales spécifiques, devint une théorie générale d'après laquelle chaque fonction du cerveau n'a qu'un emplacement câblé, idée résumée par la formule « une fonction, un emplacement », ce qui signifiait que si une zone était lésée, le cerveau ne pouvait compenser la carence qui en résultait en se réorganisant.
Ce fut le début d'une période sombre pour la plasticité. Toute exception au dogme « une fonction, un emplacement » était ignorée. En 1868, un autre praticien français, Jules Cotard, se pencha sur le cas de plusieurs enfants atteints d'une grave maladie cérébrale qui se manifestait par le dépérissement de l'hémisphère gauche, y compris la zone de Broca. Malgré cela, ces enfants arrivaient à parler normalement, ce qui signifiait que même si la fonction langagière était localisée dans l'hémisphère gauche, comme le soutenait Broca, le cerveau était suffisamment plastique pour se réorganiser si nécessaire. En 1876, l'Allemand Otto Soltmann pratiqua l'ablation du cortex moteur, la partie du cerveau en charge de la motricité, sur des chiens et lapins nouveau-nés, et constata qu'ils étaient encore capables de bouger. Ces observations furent submergées par la vague d'enthousiasme localisationniste.
LOCUS COERULEUS
Le locus cœruleus ou LC (la tache bleue, en latin) est un noyau sous-cortical du cerveau, situé dans le tronc cérébral et entretenant d'étroites relations avec l'amygdale, il a été découvert et décrit pour la première fois par le médecin et anatomiste français Félix Vicq d'Azyr au milieu du XVIIIe siècle.
Ce noyau constitue une structure noradrénergique (relatif à la noradrénaline) et cholinergique (relatif à l'acétylcholine). C'est dans cette région que partent la grande majorité des neurones qui utilisent la noradrénaline comme neurotransmetteur1. Sa couleur bleue est due a la présence de granules de mélanine, ce qui lui a aussi valu le nom de « noyau pigmentosus pontis » (noyau très pigmenté du pont).
Par son très grand nombre de connexions, le locus coeruleus est impliqué dans de nombreux effets naturels, tels que :
Par ses projections d'axones vers des régions tel que l'amygdale, l'hippocampe, le septum, le cortex, le tronc cérébral ou encore la formation réticulée, le locus cœruleus s'est vu associé aux troubles de paniques. Il a en effet été prouvé que la stimulation de cette zone provoquait un état d'anxiété chez l'animal et que, inversement, des substances tranquillisantes tel que l'alcool, les benzodiazépines ou les opiacés diminuaient son activité1.
Ses connexions avec les noyaux du raphé (connus pour leur rôle dans l'endormissement), les noyaux paraventriculaires de l'hypothalamus (rôle dans le contrôle de la posthypophyse), le neocortex cérébral (ayant un rôle majeur dans les processus d'attention et d'éveil) et l'intervention systématique du systeme noradrenergique lors de l'éveil, font du locus cœruleus un acteur indispensable au bon fonctionnement de cette « boucle veille-sommeil »
L'ensemble de ces trois aires correspond à la zone principale réceptrice au sens du toucher. Elle constitue une représentation du corps que l'on désigne parfois sous le nom d'homonculus sensitif : une projection inversée du corps, visible dans l'organisation du cortex. Jambes et tronc sont représentés sur la ligne médiane, bras et mains vers le milieu du gyrus, tandis que le visage est représenté dans la partie basse du gyrus post central. Certaines parties de cortex sont plus grandes en proportion par rapport à la taille relative au corps, des zones qu'elles représentent. Ainsi, par exemple, mains et lèvres ont leurs projections sur une importante partie de cortex par rapport aux pieds et aux jambes. Ceci est à mettre en lien avec le nombre de récepteurs tactiles, plus important dans les mains et les lèvres que dans d'autres parties du corps. On note ainsi une correspondance assez étroite entre la sensibilité au toucher des organes et la portion de cortex somatosensoriel qui les représente.
Chacune des aires 3, 1 et 2 comporte des neurones qui projettent leur axone dans les aires somatosensorielles secondaires (Aires 5 et 7).
Les lésions touchant le cortex somatosensoriel provoquent des déficits cognitifs caractéristiques de la sensibilité : astéréognosie, abarognosie, défaillances de la proprioception ou déficits de la sensibilité fine... Certaines études de cas suggèrent qu'elles pourraient affecter d'autres domaines sensitifs tels que la nociception ou la thermoréception, ou d'autres caractéristiques du toucher. Cependant, ces dernières sont également traitées par d'autres structures telles que le gyrus cingulaire et le cortex insulaire, aussi ne sont-elles pas toujours visibles ni présentes. Si les lésions se localisent dans l'hémisphère non dominant, elles entrainent classiquement le phénomène d'héminégligence.
Comme le montrent Penfield et al., cette région, tout comme le cortex somatosensoriel primaire, présenteune représentation du corps dans son organisation, on retrouve ainsi les projections motrices des jambes et du tronc, sur le dessus de la structure. Bras et mains vers le milieu, et visage vers le bas de la figure. Cette représentation est généralement nommée Homonculus moteur.
Cette région est particulièrement étendue chez l'homme mais son rôle est encore mal compris. Les hypothèses les plus intéressantes suggèrent que ce cortex joue un rôle régulant d'autres activités cérébrales, exerçant ainsi une "supervision" permettant la planification et le contrôle de l'action et des fonctions mentales, de même que leur coordination.
le processus plus complexe de compréhension du langage se déroule en fait dans les aires préfrontales, à mi-chemin entre les aires 45 et 11 de Brodmann
Aires 11 et 12. (Région du cortex préfrontal) impliquée dans la mémoire des traits et caractéristiques des objets. Gyrus orbital.
Le cortex orbitofrontal (aires de Brodmann 12 et 13) jouerait un rôle dans les fonctions nécessitant un contrôle frontal du système limbique telles que l’inhibition, le codage de la valeur motivationnelle d’un objet ou d’un stimulus, la prise de décisions et le contrôle de l’action basés sur la récompense, le contrôle de l’impulsion et des interférences, l’humeur, et la personnalité/le comportement social (Bechara et al., 2000; Rolls, 2000 ).
Au moins un groupe de recherche (1) pense avoir trouvé une aire homologue chez l'homme, présentant des caractéristiques anatomiques semblables, notamment sa localisation.
L'aire 15 reçoit des informations en provenance des nerfs de Hering, donc de l'information issue des récepteurs situés dans les carotides (barocepteurs sensibles à lapression artérielle et chimio-récepteurs sensibles auxconcentrations sanguines de divers composés). L'aire 15 est notamment suractivée lors d'attaques de paniques.
Le cortex visuel primaire est subdivisé en 6 couches fonctionnelles distinctes. L'une des plus importantes est la 4ème couche qui reçoit la majorité de l'information issue des corps génouillés latéraux (noyau géniculé latéral). Cette 4ème couche se subdivise elle même en 4 sous-couches distinctes, 4a, 4b, 4c-alpha (recevant l'apport des voiesmagnocellulaires) et 4c-bêta (voiesparvocellulaires).
L'aire 17/V1 représente une carte inversée du champ visuel : ainsi, une information issue du champ visuel bas-droit est traitée majoritairement par la partie haut-gauche du cortex visuel primaire. On nomme rétinotopie cette carte virtuelle. Cette représentation (en fait, de la rétine plutôt que du champ visuel) est très précise, même les angles morts y sont représentés. Cette rétinotopie se trouve chez la plupart des animaux ayant un cortex virtuel primaire. Lorsqu'existe une fovéa, l'aire correspondante est particulièrement grande en rapport au reste de la rétine.
David Hubel et Torsten Wiesel ont montré l'organisation en colonne des cellules du cortex visuel primaire. De telles propriétés seraient à même d'expliquer la sensibilité à l'orientation d'un stimulus visuel (vertical, oblique...) et la sensation de profondeur due au couplage des données issues des deux cortex visuels. Néanmoins, l'interprétation de l'organisation en colonne reste un sujet vif et ouvert de l'actualité scientifique contemporaine.
Les premiers neurones qui réagissent à l'information visuelle semblent traiter l'information pour en tirer des propriétés telles que l'orientation, la vitesse, le mouvement et sa direction
Un peu plus tard, après 100 ms de délai, d'autres neurones sont plus sensibles à l'organisation globale de la scène, mais il est probable qu'un premier traitement des aires visuelles supérieures rétroagisse pour permettre ce traitement.
L'information visuelle n'est pas réellement traitée en terme de propriétés de luminance ou de couleur de la scène visuelle, mais plutôt en terme de contraste : quelques neurones vont coder ces informations, tandis qu'une majorité réagira aux lignes de fractures de couleurs ou de luminance.
C'est la seconde aire majeure du cortex visuel, située entre l'aire visuelle primaire et le cortex visuel associatif, d'où son nom decortex visuel secondaire. Elle entretient de fortes connexions avec ses voisines, projetant de nombreux axones vers les aires visuelles associatives (V3, V4, V5), mais également en feedback vers l'aire visuelle primaire, dont elle reçoit la majorité des connexions.
Comme l'aire V1, elle est subdivisées en quatre parties distinctes, dorsales et ventrales, pour chacun des hémisphères. V2 partage de nombreuses propriétés avec l'aire visuelle primaire (par exemple, l'excitation liée à l'orientation des stimuli visuels, la sensibilité au contraste), mais possède également des propriétés propres, qui lui permettent vraisemblablement de traiter l'information visuelle davantage en profondeur (orientation d'un contour, appartenance d'un stimulus à une figure ou à un fond...). Elle est également un peu sensible à la régulation exercée par l'attention, plus que l'aire visuelle primaire mais moins que les aires visuelles associatives.
Des études menées sur le chat suggèrent que cette zone présente une sensibilité au mouvement d'objets délimités, permettant ainsi de différencier et d'accrocher visuellement un objet par rapport à la scène visuelle. Néanmoins, d'autres études menées sur le singe donnent des résultats variables. Le consensus actuel suggère alors la présence d'une collection hétérogène de représentations incomplètes de la scène visuelle, dans cette zone.
Chez l'homme, l'aire 19 contient les quatre aires fonctionnelles V3, V4, V5 et V6. L'imagerie fonctionnelle suggère l'existence de cartes rétinotopiques dans l'aire 19, en lien avec les aires visuelles primaires et secondaires, mais également avec certaines zones pariétales postérieures et temporales inférieures.
L'aire 19 reçoit également directement des informations dupulvinar et du colliculus supérieur, concourant alors au phénomène devision aveugle lorsque les aires primaires ou secondaires sont lésées. Chez les patients aveugles de naissance, cette zone est activée par des stimuli somatosensoriels.
Face à ces constatations, la communauté scientifique estime que l'aire 19 est une plaque tournantes des informations sensorielles multimodales, et un point de différenciation de deux types d'information visuelle : le "quoi" (discrimination visuelle) et le "où" (localisation spatiale). La région dorsale semble se spécialiser dans le mouvement d'objets visuels, tandis que la région ventrale s'attache à la reconnaissance visuelle.
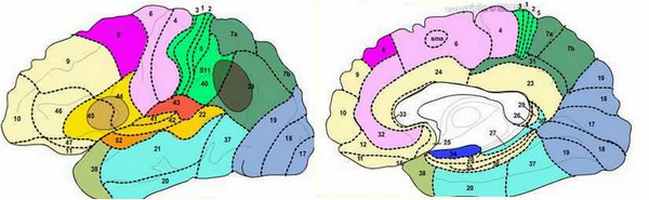
Des études réalisées sur les macaques (Macaca Mulatta) ont amené à subdiviser l'aire 23 en trois zones, dont l'une est encore subdivisible en deux parties, l'une, postéro ventral (pv23b) de cette aire 23, recevant les projections thalamiques des noyaux antérieurs, et l'autre, dorsale (d23b), dont les connexions avec ces noyaux sont beaucoup moins nombreuses. Fonction à déterminer.
A l'origine partie intégrante de l'aire 24, cette aire est dissociée par Brodmann dans ses travaux ultérieurs (1909).
Cette région semble impliquée dans le traitement olfactif et dans la gestion de l'humeur. Ainsi, une étude réalisée par Mayberg et al (2005) montrait l'amélioration de l'humeur de dépressifs résistant aux traitements classiques, suite à la stimulation cérébrale profonde de cette zone.
Cette structure occupe une petite partie du cerveau chez l'homme et les primates proches, par rapport aux autres animaux, surtout les insectes
Sa cytoarchitectonie est particulière : elle est subdivisible en au moins 7 aires secondaires, dont l'une, TG, est spécifique à l'humain. On soupçonne cette aire TG d'être impliquée dans des processus perceptuels de haut-niveau, ainsi que dans des processus également de haut niveau, de réponses émotionnelles et sociales.
Cette région est l'une des premières touchées lors d'une maladie d'Alzheimer, et l'une des premières concernées par les épilepsies temporales.
Cette région est subdivisible en 5 zones distinctes parmi lesquelles la zone PF connue pour être une partie du système des neurones miroir, active lorsque l'homme imite un pair.
La partie de l'aire 40 située dans le gyrus supramarginal (lobe pariétal inférieur) est impliquée dans la lecture, tant pour lasémantique (signification) que laphonologie.
Le fonctionnement amygdalien est perturbé lors d’un épisode dépressif. En effet, on observe une augmentation de son volume chez des sujets pris en charge pour un premier épisode dépressif. Les études d’imagerie fonctionnelle mettent en évidence une réactivité amygdalienne plus importante chez les sujets déprimés au cours de la présentation de stimuli à forte charge émotionnelle, confirmant ainsi le rôle de cette structure dans la régulation des émotions. La perturbation du fonctionnement amygdalien est ainsi responsable des symptômes que l’on retrouve chez les patients déprimés : une agitation anxieuse, une irritabilité, une hypervigilance, des réactions de sidération fréquentes.
Quel est ce prodige ? Existe-t-il dans le cortex visuel des neurones spécialisés dans la détection et la localisation des araignées et des visages en colère ? En fait, le visage en colère ne saute pas vraiment aux yeux comme peut le faire un coquelicot dans un champ de blé au printemps ; il ne s'agit donc pas vraiment d'un effet Pop Out de Treisman utilisant des détecteurs spécialisés dans les cortex visuel et auditif. Quand le nombre d'objets entourant le visage augmente, la recherche prend plus de temps, au contraire du mécanisme Pop Out. Il faut donc chercher ailleurs le responsable de ce mode de recherche accéléré. À l'heure actuelle, le principal suspect semble être une petite structure cérébrale en forme d'amande appelée amygdale, qui n'a rien à voir avec les organes de la gorge que l'on enlève parfois chez les enfants. L'amygdale est nichée confortablement à l'avant du lobe temporal, et fait partie du système limbique, en charge des émotions. Comme il y a deux hémisphères cérébraux, il y a d'ailleurs deux amygdales, une de chaque côté.
Publiée en ligne dans Nature Neuroscience, une étude expérimentale américaine montre qu’une partie de notre cerveau, l’amygdale droite, est tout spécialement activée à la vue d’un animal, quel qu’il soit : un héritage de notre passé dans la nature sauvage, sans doute.
Des chercheurs de l’Institut de technologie de Californie (Caltech) et de l’Université de Californie (UCLA) ont travaillé avec 41 patients atteints d'épilepsie et traités au Centre médical Ronald Reagan. En utilisant les électrodes déjà en place dans les cerveaux de ces personnes, l'équipe a enregistré les réponses du complexe amygdalien – une zone située profondément dans le lobe temporal du cerveau – à certains stimuli visuels : images d’animaux, de gens, d’objets ou d’éléments du paysage. Cette expérience leur a alors révélé un mécanisme étonnant dans cette structure de notre cerveau.
"Notre étude montre que les neurones de l'amygdale humaine répondent préférentiellement à des photos d'animaux : (…) nous avons observé une plus grande activité dans les cellules lorsque les patients regardaient des chats ou des serpents que lorsqu’ils regardaient des bâtiments ou des gens. Cette ‘préférence’ s'étend aux animaux mignons aussi bien qu’aux animaux laids ou dangereux, et semble être indépendante du contenu émotionnel des images. De façon remarquable, nous n’avons trouvé ce comportement de réponse que dans l’amygdale droite, et non dans la gauche", explique ainsi Florian Môrmann, auteur principal de l’étude.
Une asymétrie hémisphérique qui corrobore l'idée que, dès le début de l'évolution des vertébrés, l'hémisphère droit s'est spécialisé dans le traitement des stimuli inattendus et biologiquement pertinents. "En termes d'évolution du cerveau, l'amygdale est une structure très ancienne, et tout au long de notre histoire biologique, les animaux - prédateurs ou proies – ont constitué une classe très pertinente de stimuli…", conclut le chercheur.
1 - La neuropathologie anxieuse
2 - L’exercice régulier : un remède contre l’anxiété
Les neurotransmetteurs de l'anxiété-1
Les neurotransmetteurs de l'anxiété-2
Les neurotransmetteurs de l'anxiété-3Les marques de l'anxiété dans les circuits de l'amygdale-1
Les marques de l'anxiété dans les circuits de l'amygdale-2
Les marques de l'anxiété dans les circuits de l'amygdale-3
Anormalités cérabrales liées aux troubles anxieux-1
Anormalités cérabrales liées aux troubles anxieux-2
Anormalités cérabrales liées aux troubles anxieux-3
L'accident ischémique transitoire ou AIT est un déficit neurologique localisé, survenant brutalement et disparaissant complètement en moins de 24 heures.
Il est secondaire à l'occlusion d'une artère cérébrale ou médullaire.
Lorsque le sang n'arrive plus dans le tissu cérébral, il se crée un infarctus ou un ramollissement.
Les causes sont nombreuses :
Les plaques d'athérome (athérosclérose) sur les vaisseaux cervico-céphaliques provoquant un rétrécissement du diamètre des vaisseaux avec risque de sténose pouvant déclencher un accident ischémique transitoire ;
Les cardiopathies emboliques pouvant provoquer une embolie cérébrale : rétrécissement mitral , fibrillation auriculaire et autres troubles du rythme, endocardite, thyréotoxicose. Elles suivent le trajet artériel et touchent principalement le territoire sylvien gauche (vascularisé par l'artère sylvienne gauche) ;
Les dissections des artères cervicales et intracrâniennes qui sont plus fréquentes chez le sujet jeune.
Les accidents transitoires se traduisent par un épisode neurologique passager totalement régressif dont la durée est de quelques secondes à quelques minutes. Il peut durer parfois quelques heures. Les symptômes apparaissent et disparaissent brutalement.
En fonction du territoire atteint, il peut s'agir d'une cécité d'un oeil transitoire, d'une amputation du champ visuel, de troubles sensitifs ou moteurs localisés, d'aphasie, de troubles cérébelleux, etc.
L'AIT est un signal d'alarme. Il peut prévenir de l'imminence d'un accident vasculaire permanent et doit faire déclencher une enquête. Cet accident vasculaire permanent peut survenir dans les heures qui suivent un AIT ou bien plusieurs années plus tard ou même jamais. Rien ne permet de le prévoir.
vidéos sur l'AVC :
AVC/1 -
AVC/2
- AVC/3
- AVC/4
1 - la peau
2 - la langue
3 - le nez
4 - les yeux
5 - les oreilles
La peau comporte différents types de capteurs :
1 - des fibres sensitives qui se terminent librement dans l'épiderme et qui transmettent la douleur
2 - des fibres coiffées de capteurs ressemblant à des bulbes d'oignons nommés "Corpuscules de Pacini"
3 - des fibres coiffées d'autres capteurs nommés les "Corpuscules de Meissner" responsables du toucher (les "Corpuscules de Krause" captent les variations thermiques du froid et les "Corpuscules de Ruffini captent le chaud)
4 - La base des poils riche en terminaisons nerveuses (amas de neurones en forme de tonneau)
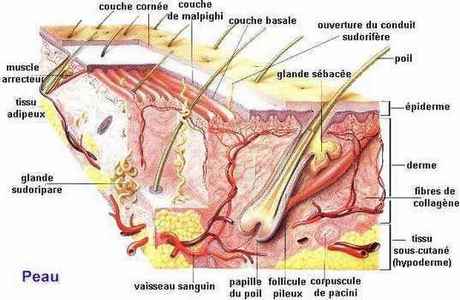
1 - la voie lemniscale (lemniscus = ruban) qui est rapide et ponctuelle (3 neurones cablés en série). Les trois neurones se relaient jusqu'au cortex cérébral et plus exactement dans le lobre pariétal opposé (les voies sensitives "croisent" comme les voies motrices), immédiatement en arrière de la scissure de Rolando
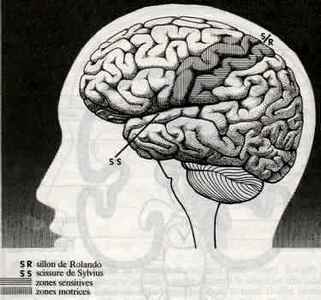
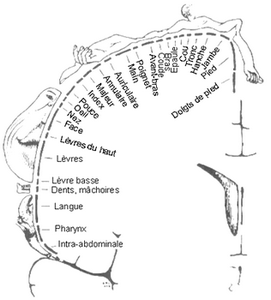
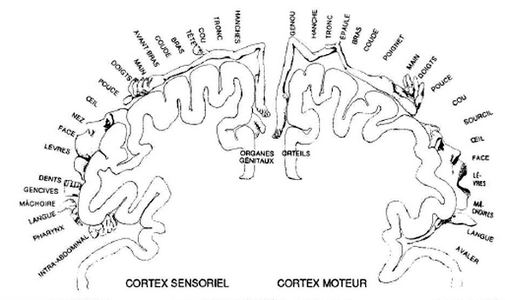
La peau est le plus grand organe sensoriel au regard de sa surface et de sa masse : chez l'adulte, environ 2 m2 pour 5 kg. Les récepteurs au toucher ne se trouvent pas à la surface de la peau, comme on pourrait le croire, car une couche insensible recouvre tout le corps. La plus grande concentration des récepteurs se trouve dans une deuxième couche plus profonde. Les zones les plus denses en récepteurs, comme le bout des doigts ou les lèvres, sont aussi les plus sensibles. La sensibilité à la douleur varie selon les parties du corps. Par exemple, une très légère pression provoque une douleur dans l'oeil, alors qu'il en faut une beaucoup plus forte pour se faire mal à la paume de la main. Quand un des récepteurs est stimulé, il envoie un message au cerveau.
Le toucher est un autre mécanisme de défense du corps. La douleur prévient d'une blessure ou d'un danger. Si nous sentons quelque chose de très chaud, nous apprenons à retirer notre main. Si nous éprouvons une sensation de coupure, nous interrompons le geste commencé. Certaines maladies affectent cette sensation de douleur et des personnes souffrant de la lèpre ne sentent ni la douleur ni la chaleur, ce qui les rend incapables de réaliser s'ils courent le risque de se brûler ou de se blesser.
Des médicaments comme la morphine peuvent aussi atténuer la sensation de la douleur. De plus, il arrive que le cerveau bloque la douleur en libérant des substances chimiques qui s'appellent des endorphines. En condition de stress extrême, comme lors d'une compétition sportive, les athlètes peuvent ignorer la douleur immédiate et ne la ressentir qu'une fois la compétition achevée. Le même phénomène peut se produire dans une guerre, où certains soldats évacuent la douleur pour sauver l'un des leurs...
Le comportement des athlètes et des soldats illustre aussi le fait que les individus perçoivent la douleur de manière différente. Une personne pourra ne prendre qu'un cachet d'aspirine ou un médicament en vente libre quand un autre, pour contrer la même douleur, aura besoin d'antalgiques puissants qu'il ne pourra se procurer que sur ordonnance. C'est particulièrement vrai des personnes souffrant d'affections chroniques, comme le mal de dos.
Les gens ont aussi des réponses différentes à la douleur selon les situations. Imaginons que vous accrochez un tableau au mur et que vous vous tapez sur le doigt avec un marteau. Ce type d'accident fait très mal et vous aurez le pouce enflé pendant 48 ou 72 heures. Si vous avez une réunion de famille qui vous ennuie le lendemain, votre douleur sera un bon prétexte pour annuler. Mais si votre meilleur ami vous offre une place pour le match de l'année au PSG, il y a de grandes chances que vous oubliiez votre mal !
Notons que le cerveau lui-même ne sent rien, parce qu'il n'a aucun récepteur tactile. Les neurochirurgiens peuvent donc pratiquer des opérations du cerveau sur des patients éveillés, ce qui est important quand l'intervention touche des zones aussi décisives que le contrôle de la parole ou du mouvement des bras et des jambes.
Plus de détail sur la peau et les phanères
Le sens du goût est étroitement lié à l'odorat et on compte quatre goûts fondamentaux : l'amer, le sucré, le salé et l'acide, qui peuvent se combiner pour former plus de 10 000 parfums. Les papilles gustatives sont les petites bosses disposées à la surface de la langue dans lesquelles sont regroupés les bourgeons du goût. Les cellules sensorielles sont, quant à elles, concentrées dans ces bourgeons du goût.
Les cellules sensibles à un goût particulier ne sont pas réparties uniformément sur toute la langue. Les cellules sensibles au sucré sont concentrées au bout de la langue. Sur les côtés il y a les cellusles sensibles aux substances salées et acides. Encore plus au fond, c'est la zone qui détecte le goût amer. Enfin, toutes les papilles ne sont pas sur la langue. Il y en a aussi dans la gorge, à l'intérieur des joues et sur le palais.
On trouve la répartition des papilles gustatives dans tous les livres sur les sens, mais des recherches plus récentes ont conduit à penser que cette description n'est pas assez précise. Il est possible en fait que chaque cellule d'une papille gustative réponde individuellement à tous les goûts et que la distinction se fasse au moment où le message des récepteurs de la langue arrive au cerveau.
Comme pour les cellules impliquées dans les autres sens, une molécule spécifique paraît associée à un récepteur particulier, un peu comme une clé et sa serrure. Dans le cas du goût, les molécules de nourriture ont la clé d'une papille gustative précise qui, à son tour, produit un signal électrique transmis au système limbique et au cortex cérébral.
Les centres moteurs du cerveau donnent l'ordre aux muscles de soulever la fourchette jusqu'à la bouche, de mâcher et d'avaler. Ils ordonnent aussi aux glandes de saliver. Le système limbique est le centre du plaisir, c'est lui qui vous donne le sentiment d'aimer ou de détester ce que vous goûtez. Le cortex frontal, ensuite, envoie un message au centre de la parole qui vous fait dire : « C'est délicieux ! » ou « Beurk ! » (comme font presque tous les enfants devant un plat de légumes !).
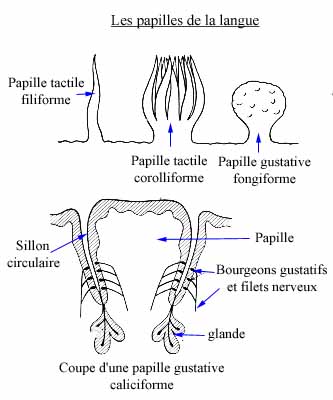
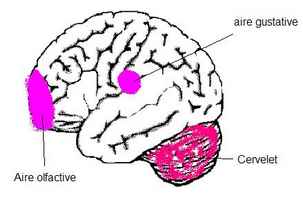
Comme la vision et l'audition, le sens du goût décline avec l'âge. On dispose de 10 000 papilles gustatives au début de notre vie. À 60 ans, on en a moins de 7 000, ce qui explique que les personnes âgées ont un sens gustatif plus limité et ont tendance à aimer les plats épicés.
La muqueuse compte environ 1 million de cellules spécialisées (neurones olfactifs) équipés de cils long de 3 à 5 miclbe olfactif C'est donc en réalité le "cerveau qui goute" . Ce mécanisme est lié aux échanges de sodium et potassium dans les papilles gustatives. Autrement dit, c'est notre mémoire qui nous permet de reconnaître le goût des aliments.
De plus, le cerveau permet de transformer cette sensation en émotrons qui baignent dans un liquide épais recouvrant la muqueuse. C'est par l'intermédiaire de ce mucus que le neurorécepteur olfactif est en contact avec le milieu extérieur. Seules les molécules volatiles, piégées et solubles dans le mucus seront perçues. En atteignant les cils elles déclenchent un stimulus nerveux.
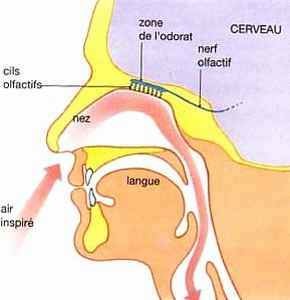
Comment fait exactement la personne qui identifie une odeur ? Une grande partie du processus reste mystérieuse, mais on a pu reconstituer quelques éléments.
Les narines conduisent à une cavité située au-dessus et derrière le palais. Chaque cavité nasale est recouverte d'un petit bout de tissu plus petit que l'ongle et qui contient environ 5 millions de cellules olfactives. Il s'agit de la muqueuse olfactive. De même que celles de la cochlée, ces cellules ont de minuscules cils (6 à 8 chacune), qui détectent les substances chimiques à l'origine de l'odeur.
Comme pour l'audition, les chiens ont un meilleur odorat que les humains (200 millions de cellules réceptrices de l'odorat pour le chien contre 40 millions pour nous). La truffe d'un chien est aussi mieux équipée pour sentir les odeurs, parce qu'elle est en général grande et humide. Les amis des bêtes trouvent cette truffe adorable, mais elle est surtout « odorable », parce qu'elle recueille et absorbe les particules d'odeurs d'une façon beaucoup plus efficace que ne le fait notre petit nez tout sec.
L'information sur les odeurs qui pénètrent dans le nez est instantanément traduite en un signal nerveux, lequel parcourt ensuite environ 2,5 cm pour atteindre l'aire du cerveau qui l'interprète.
On soupçonne - sans en être vraiment sûr - que le sens de l'odorat fonctionne un peu comme l'aptitude
à distinguer les couleurs. Rappelez-vous que les cellules de la rétine ne peuvent recevoir les ondes lumineuses que de trois couleurs différentes et que c'est la combinaison de ces couleurs qui donne les nuances de l'arc-en-ciel. Les scientifiques ont identifié sept odeurs de base (camphrée, musquée, florale, mentholée, éthérée, piquante et putride) et font l'hypothèse que toute la gamme des odeurs serait la combinaison de ces sept odeurs.
Cette hypothèse signifie qu'il n'y a probablement pas, dans le nez, de récepteur spécifique permettant d'identifier une odeur de rose ou putois. Pourtant, le cerveau est capable de distinguer les deux. Les axones des neurones de la muqueuse olfactive passent à travers des stations relais (appelées bulbes olfactifs). L'information olfactive parvient ensuite au thalamus, qui lui-même est connecté au cortex frontal. À ce niveau, les signaux olfactifs sont comparés à ceux qui sont en mémoire pour la reconnaissance des odeurs. D'autres neurones de la muqueuse olfactive transmettent des informations au système limbique, qui est impliqué dans la motivation, l'émotion et certains types de mémoire. Le fait que l'interprétation de l'odeur soit réalisée précisément par cette partie du cerveau aide à expliquer l'association que font certaines personnes entre des odeurs et des souvenirs ou des sentiments particuliers. Rappelez-vous qu'un certain Marcel Proust a pu écrire des pages et des pages sur l'odeur d'une petite madeleine qui lui fait revivre une scène de son enfance... Et nous y avons gagné un chef-d'oeuvre de la littérature !
Les gens peuvent détecter des milliers d'odeurs. Comment se fait-il qu'ils ne se laissent pas submerger par les senteurs de toutes sortes ? La réponse est simple : le cerveau filtre les odeurs comme il filtre les sons. De même qu'on peut s'habituer à un son, de même on se désensibilise à certaines odeurs. Mais il y a des odeurs auxquelles on ne s'habitue jamais.
Plus de détail sur l'olfaction
Les yeux sont des petites sphères contenant une gelée transparente qui les maintient bien rondes. Chaque globe oculaire a un diamètre de 2,5 cm, une masse d'environ 7 grammes et un volume d'environ 6,5 cm3. Le blanc de l'oeil, c'est la sclérotique, qui est une enveloppe de protection. Six petits muscles striés assurent le mouvement du globe oculaire, droit devant, en haut, en bas, sur les côtés.
L'oeil est recouvert par la cornée, une membrane solide et transparente. Ce qui donne sa couleur à l'oeil (bleu, vert, gris ou marron), c'est la présence de pigments de mélanine dans l'iris (arc-en-ciel en grec). La pupille est ce trou noir au centre de l'iris qui permet à la lumière d'entrer dans l'oeil.
L'oeil humain est souvent comparé à un appareil photo parce que les fonctions de leurs composants sont les mêmes. La pupille, comme le diaphragme de l'appareil photo, contrôle l'intensité de la lumière qui pénètre dans l'ceil : grande ouverture quand la lumière est faible, petite ouverture quand la lumière est vive. Le cristallin de l'ceil, comme un objectif, fait la mise au point et focalise la lumière sur la rétine. Quand une personne regarde des objets éloignés, les muscles autour du cristallin se relâchent, de sorte que ce dernier rétrécit. Les muscles se contractent et le cristallin s'agrandit quand la personne regarde des objets proches. Le fond de l'oeil est tapissé de cellules photosensibles qui changent quand elles sont exposées à la lumière, comme la pellicule photo. Cette couche de cellules s'appelle la rétine.
La rétine dispose de deux types de cellules photosensibles : les cônes et les bâtonnets. Les cônes fonctionnent mieux si la lumière est vive; ils aident à voir la clarté et la couleur du stimulus. Les 7 millions de cônes sont sensibles aux trois couleurs : le rouge, le bleu et le vert.
Plus nombreux que les cônes (125 millions), les bâtonnets ont une très grande sensibilité à la lumière. Les bâtonnets et les cônes convertissent la lumière en signaux nerveux transmis au cerveau.
L'image qui se forme sur la rétine est inversée : cela s'explique par la manière dont les deux lentilles de l'oeil (la cornée et le cristallin) font converger les rayons lumineux qui entrent dans l'oeil. L'information visuelle voyage par le nerf optique jusqu'au chiasme optique. À ce niveau, les fibres de la rétine nasale de chaque oeil vont bifurquer vers la moitié opposée du cerveau. Les fibres de la rétine temporale (côté oreille) ne bifurquent pas. Le principal centre de traitement des informations transmises par ces fibres est le centre visuel du cortex, dans le lobe occipital.
De là, des signaux plus spécifiques sont transférés dans d'autres régions du cerveau. Les savants ont longtemps cru que les objets envoient des codes visuels qui s'impriment sur la rétine et que le cortex visuel décode le message et le traduit en une image complète. Plus tard, au cours du xxe siècle, de nouvelles recherches ont montré que les impulsions neuronales venant des yeux sont décomposées en plusieurs éléments. Ainsi, le message relatif à la couleur des objets va vers une certaine aire, les signaux sur les lignes horizontales sont dirigés vers une autre zone et les transmissions relatives à la position de la tête dans l'espace sont traitées dans une troisième aire. Pendant ce temps, les signaux associés à la perception et à la reconnaissance des objets vont vers les centres supérieurs du cortex, où ils sont analysés et comparés avec les souvenirs emmagasinés. Le cerveau rectifie aussi l'orientation de l'image et la remet à l'endroit.
Pour avoir une idée du fonctionnement du système visuel, imaginez que vous êtes en train de chasser dans les bois et que vous voyez un animal qui bouge dans les buissons. Grâce aux nerfs crâniens, vos yeux se concentrent et se fixent sur l'endroit où cela bouge. Les nerfs transmettent aussi les messages du cerveau qui ordonne à vos muscles de bouger la tête pour suivre le mouvement des buissons. Le cerveau calcule la taille et la vitesse de l'animal et compare les images qui lui arrivent par les yeux avec les souvenirs de formes, de couleurs et de mouvements similaires. La possibilité que l'animal soit un ours stimule les ganglions de la base, le cervelet et le cortex moteur pour augmenter le niveau de vigilance du corps. Ordre est donné au coeur de battre plus vite, aux poumons d'inspirer plus d'oxygène. Quand l'animal sort des buissons et que le cerveau reconnaît l'image d'un cerf inoffensif, les muscles reçoivent le message de se relâcher.
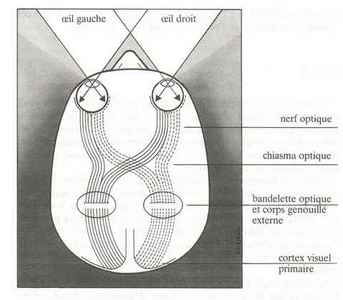
Les humains ont deux yeux qui bougent ensemble (au contraire, les yeux d'un caméléon peuvent bouger séparément, ce qui fait qu'il peut voir dans deux directions simultanément). Cette vision binoculaire a plus d'avantages qu'une vision monoculaire. Car deux yeux permettent d'avoir un plus grand panorama qu'un seul oeil, avec un champ visuel de 200 degrés environ. Le champ visuel commun à l'oeil droit et à l'oeil gauche est un peu plus restreint (140 degrés) et c'est leur chevauchement qui définit notre vision en 3-D. Le champ visuel nous aide à juger des distances avec précision.
Tout le monde a passé au moins une fois le test de Snellen, qui consite à lire des lignes de lettres, d'abord très grosses, puis de plus en plus petites. Une note sur 10 est donnée à l'issue du test. Mais savez-vous ce que cette note signifie réellement ? La graduation de l'acuité visuelle s'écrit sous forme de fraction. Le nombre supérieur de la fraction désigne la distance à laquelle la lecture des plus grosses lettres est possible pour une personne ayant une acuité visuelle normale et le nombre inférieur désigne la distance à laquelle la personne examinée réussie à lire cette lettre. On considère que la vision normale s'effectue à 6 mètres et elle est notée 6/6 (ou 10/10 en notation décimale). Une personne avec une acuité de 1/10 doit s'approcher à 6 mètres pour voir une lettre qu'une personne dotée d'une vision normale peut lire à 60 mètres. Une personne avec une acuité visuelle de 20/10 peut voir à 6 mètres une lettre qu'une personne avec une vision normale lira à 3 mètres.
Outre le test de Snellen, vous avez probablement passé le test Ishihara pour le repérage du daltonisme, dans lequel il vous est demandé de reconnaître un chiffre à l'intérieur d'un disque de points colorés. La personne ayant une vision normale peut lire le chiffre à retrouver tandis qu'une personne daltonienne verra un chiffre complètement différent. Un daltonien ne peut pas voir la gamme normale des couleurs et certaines peuvent lui apparaître grises. La forme la plus commune de daltonisme est le daltonisme rouge-vert, qui se caractérise par une difficulté à distinguer les rouges, les verts et les marron. Le daltonisme touche beaucoup plus d'hommes que de femmes, parce que l'anomalie à l'origine du daltonisme concerne les chromosomes sexuels X et Y.
Autant le daltonisme est incurable, autant les lunettes peuvent corriger les problèmes de vision décelés par le test de Snellen ou d'autres tests. Pour avoir une vision nette et claire, les lentilles de chaque oeil (cornée et cristallin) doivent focaliser les rayons lumineux sur un point précis de la rétine, la fovéa. Ce point est située dans le prolongement de l'axe optique de l'ceil. Quand les lentilles d'un oeil ne font pas converger correctement les rayons lumineux sur la rétine, cela produit des images floues.
Par exemple, des objets lointains paraissent flous à un myope parce que les lentilles de l'oeil font converger les rayons lumineux en avant de la rétine. L'image qui impressionne la rétine est donc plus étalée et plus floue. Des lunettes à verres concaves permettent la focalisation des rayons lumineux sur la bonne zone de la rétine et rendent ainsi l'image plus claire. Lentilles et lunettes ont longtemps permis de pallier ce défaut de vision, mais il est aujourd'hui possible d'opérer, entre autres grâce à la technique du Lasik (laser).
Même la meilleure vue est susceptible de se détériorer. L'élasticité du cristallin tend à diminuer avec le temps, ce qui rend plus difficile la mise au point. La cornée peut aussi changer de forme et perdre de sa transparence, ce qui donne une image déformée et floue. En France, plus de la moitié des personnes de 65 ans et plus souffrent de cataracte, une opacification du cristallin qui réduit la lumière entrant dans l'oeil et la diffuse d'une manière anormale. La vision se trouble et il devient plus difficile de lire, de conduire, de bien voir. Aujourd'hui, l'opération de la cataracte est devenue assez courante et donne en règle générale de très bons résultats.
Parfois le cerveau fait des erreurs. Eh oui ! Il arrive que l'on voie quelque chose et que ce ne soit pas ce que le cerveau nous indique. Levez les yeux vers la lune par une nuit bien claire. Selon sa position dans le ciel, elle paraîtra énorme et si proche qu'on croirait pouvoir la toucher. D'autres fois, c'est une sphère minuscule et lointaine. Pourtant, la lune n'a pas changé de taille. Quand le cerveau interprète la manière dont vous voyez l'image pour la faire coïncider à un motif connu, mais qui ne correspond pas à ce que vous voyez vraiment, c'est une illusion optique.
Vous avez certainement expérimenté de nombreuses illusions d'optique. C'est le cas quand vous tracez deux segments parallèles de longueur égale et que vous ajoutez aux extrémités du premier des flèches ouvrantes et aux extrémités du second des flèches fermantes, alors les deux segments ne semblent plus être de la même longueur.
Il y a aussi beaucoup d'images que vous pouvez voir de différentes manières. Peut-être avez-vous déjà essayé ce tour dans lequel vous devez fixer un objet ou une spirale en rotation, puis regarder un mur tout blanc :vous y voyez encore l'objet ! L'artiste néerlandais Maurits Cornelis Escher (1898-1972) a créé des illusions artistiques très intéressantes, comme des bâtiments impossibles, en utilisant des motifs géométriques répétitifs ou d'autres dessins qui mettent au défi notre capacité à interpréter une image.
Plus de détail sur l'oeil
Derrière le tympan, il y a l'oreille moyenne, cavité remplie d'air qui contient les trois plus petits os du corps humain : les osselets ou, plus exactement, le marteau (malleus), l'enclume (incus) et l'étrier (stapes). Le marteau fait environ 5 à 6 mm et tient sur le bout d'un doigt.
La trompe d'Eustache est un conduit qui s'étend de l'oreille moyenne à l'arrière de la gorge. Elle permet à l'air d'entrer et de sortir pour équilibrer la pression entre l'oreille et l'extérieur. Quand la pression est inégale, par exemple lorsque vous vous trouvez dans un avion qui atterrit, le tympan ne vibre pas correctement.
Les vibrations passent le tympan, puis les osselets, zone où elles sont amplifiées 20 ou 30 fois avant d'atteindre l'oreille interne.
L'oreille interne contient un canal enroulé sur lui-même pour former un système tubulaire en colimaçon, ou cochlée. La cochlée est directement impliquée dans l'audition. Ce petit tuyau d'environ 3,5 cm s'enroule plusieurs fois en spirale comme un escargot. À l'intérieur de ses plis se trouve une membrane tapissée de plus de 20 000 cellules sensorielles, les cellules ciliées, chacune comptant entre 50 et 100 cils. Les vibrations sonores produisent des ondulations dans le liquide qui emplit la cochlée, ce qui fait bouger les membranes et rend les touffes ciliaires libres de vibrer. Les cellules ciliées transforment les ondes sonores en signaux nerveux qui sont transmis au cortex auditif dans le cerveau via le nerf cochléaire et d'autres nerfs. Cette partie du cerveau comprend 100 millions de neurones.
Chaque cellule ciliée peut émettre jusqu'à 20 000 influx nerveux par seconde, ce qui implique que 400 millions de messages relatifs au son peuvent atteindre le cerveau chaque seconde. En fait, cela ne se passe pas comme ça, car il n'y a que 30 000 fibres nerveuses pour transporter les signaux et chacune ne peut traiter que 1 000 signaux par seconde, soit 30 millions en tout. Du coup, beaucoup d'informations se perdent en route.
De même que pour les informations visuelles en provenance des yeux, la plupart des fibres nerveuses transportant le son passent du côté opposé du cerveau.
À leur arrivée, les messages sont comparés par les deux hémisphères et répertoriés dans le catalogue des sons de la mémoire. En se basant sur cette comparaison, le cerveau donne des instructions au corps pour agir ou décide au contraire de garder l'information en mémoire. Comment se passe ce processus, cela reste un mystère.
Le processus de l'audition est assez remarquable. Pensez à la façon que vous avez de filtrer certains sons. Par exemple, une mère peut dormir dans n'importe quel environnement sonore, mais si son enfant s'agite dans la pièce voisine, elle va se réveiller immédiatement. Son cerveau surveille les sons et, d'une certaine manière, elle peut sélectionner ceux qui sont importants.
La différence de perception d'un même stimulus, selon les personnes, est un autre phénomène auditif intéressant. Par exemple, un des deux auteurs de ce livre, jeune, brillant et extraordinairement beau, a le très bon goût d'aimer le rock tandis que son père, étroit d'esprit, considère que c'est juste un affreux boucan...
La vibration des molécules produit le son, qui est mesuré en décibels. Le murmure le plus doux est d'environ 10 décibels, ce qui est le plus petit son que les humains peuvent entendre. Quand le son atteint 130 décibels, il devient douloureux, mais tout bruit de plus de 90 décibels peut endommager l'audition. Les experts recommandent de mettre des boules Quiès ou toute autre protection quand le bruit excède 85 décibels, ce qui est à peine au-dessus du vacarme du trafic automobile à une heure de pointe... Une directive européenne fixe un seuil de 85 décibels comme niveau sonore maximum au travail (contre 115 décibels aux États-Unis) et restreint le temps d'exposition à un tel bruit (8 heures).
L'oreille humaine ne peut détecter qu'un champ de fréquences limité, qui se mesure en hertz. Elle peut entendre des sons qui vont de 15 à 20 000 hertz, tandis qu'un chien peut entendre des sons qui vont jusqu'à 30 000 hertz. C'est pourquoi le sifflement d'un chien est inaudible pour nous alors qu'il ne l'est pas pour nos animaux de compagnie (les chauves-souris peuvent entendre des sons
qui vont jusqu'à 100 000 hertz). La prochaine fois que vous faites les magasins pour acheter du matériel stéréo, gardez cette échelle en tête, au cas où le vendeur voudrait vous faire acquérir un appareil qui empêcherait d'entendre, parce qu'il dépasserait les fréquences que vous pouvez détecter.
L'oreille peut entendre 40 000 sons différents. Fait intéressant, détecter si le son vient de la droite ou de la gauche est plus facile que de savoir si le son vient d'en haut ou d'en bas : cela tient à la forme des oreilles, qui sont plus adaptées pour « attraper » les ondes sonores qui viennent des côtés. Les ondes sonores se propagent à une vitesse d'environ 1 200 km/h et le cerveau peut immédiatement déterminer quelle oreille reçoit le son en premier et, à partir de là, juger de la direction du son.
L'oreille est un organe très sensible, que les gens maltraitent généralement en écoutant de la musique très fort, en travaillant dans un environnement bruyant ou en tolérant la pollution sonore. Les oreilles sont résistantes, mais une exposition répétée à des sons très forts peut les endommager de manière irréversible.
Quand vous mettez le volume à fond pour écouter Led Zeppelin (et au risque de rendre vos parents fous !), vous détruisez des millions de vos neurones sensibles aux hautes fréquences. Ces pauvres bougres qui restent dans les aéroports sur le tarmac à guider les avions et qui sont constamment bombardés de bruits de décollage ont probablement des problèmes similaires de perte d'audition, même s'ils portent des protections pour les oreilles.
On estime que 5 millions de Français ont des problèmes d'audition et qu'il y a des millions de sujets à risque. En plus de la surdité, beaucoup de personnes sont affectées d'acouphènes (sifflements et bourdonnements dans les oreilles).
La surdité profonde est généralement de deux types : surdité de perception et surdité de transmission. Les personnes qui souffrent d'une surdité de perception ont un défaut physique dans l'oreille interne ou un problème de connexion entre le nerf auditif et le cerveau. Si le problème est congénital ou résulte d'un accident, il y a de fortes probabilités que la surdité est définitive. La surdité peut être partielle ou totale; de même, elle peut être partiellement corrigée par des appareils ou par des mesures plus drastiques, comme l'implant de cochlée.
Quand les vibrations sonores ne peuvent atteindre l'oreille interne, les personnes souffrent d'une surdité de transmission. Dans certains cas, le conduit auditif est bouché ou encombré de cérumen. Il suffit d'enlever le bouchon pour régler le problème. Mais des infections peuvent aussi provoquer des problèmes d'audition, par exemple quand une inflammation dans l'oreille empêche la vibration normale du tympan et des osselets. Ces états peuvent guérir d'eux-mêmes ou être traités par antibiotiques.
Une cause possible d'une perte auditive unilatérale (d'un seul côté) est la présence d'une tumeur sur le nerf acoustique à la base du cerveau (au niveau de l'angle pontocérébelleux, si vous voulez tout savoir). Cette tumeur peut être traitée en microchirurgie par une équipe comprenant un neurochirurgien et un neurologue. Ces dernières années, on traitait ces tumeurs grâce au couteau gamma (ou scalpel gamma). C'est une technique non invasive, une thérapie par faisceaux de rayons gamma.
Pour finir, rappelons que l'âge aussi contribue à une déperdition graduelle de l'audition. Donc, si vous avez l'impression de moins bien entendre et que vous n'avez plus trente ans depuis pas mal de temps déjà, inutile de vous effrayer tout seul avec des idées de tumeurs et autres maladies. Consultez votre médecin en vous disant que les années sont sans doute en train de vous jouer un tour...
L'organe de l'audition est constitué par la cochlée.
L'organe de l'équilibre (saccule, utricule et canaux semi-circulaires) enregistre les changements de position du corps, plus particulièrement de la tête.
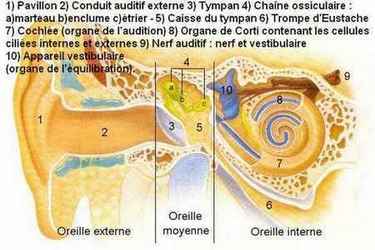
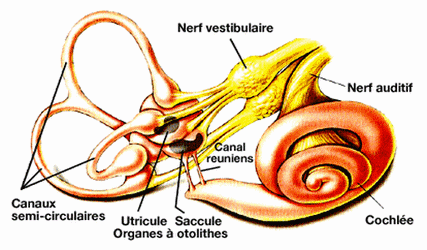
Plus de détail sur l'oreille
Le deuxième exemple appartient à la physiologie. Dennise Théodosis, dans le Laboratoire de neuro-endocrinologie de Bordeaux, a observé un curieux phénomène au cours de la lactation chez la rate. Dès l'accouchement et, plus tard, pendant toute la période où l'animal allaite ses petits, on observe, dans une région précise de l'hypothalamus, les noyaux magnocellulaires, un bouleversement total de l'architecture nerveuse qui se traduit par une disparition des cellules gliales. Les cellules nerveuses sont
alors en contact direct, leur isolant glial s'étant en quelque sorte évanoui. Or ces neurones sécrètent l'hormone qui provoque l'éjection de lait, l'ocytocine, hormone libérée dans le sang de façon périodique et paroxystique en réponse à la tétée des petits. On pense que c'est grâce à la disparition relative de la glie que se trouve favorisée l'activation paroxystique des structures nerveuses qui fabriquent l'ocytocine, une hormone qui a également son rôle dans la genèse et l'entretien du comportement maternel. On voit ici comment des mouvements de la glie, localisés à une région du cerveau, entraînent un bouleversement de la physiologie de l'animal en déclenchant tout à la fois un comportement et une sécrétion hormonale adaptés à la nouvelle situation. Après l'arrêt de la lactation, le cerveau revient à son état antérieur. C'est donc toute la manière d'être au monde d'un individu — passion maternelle — et la survie de ses petits qui sont conditionnées ici par les mouvements de quelques milliers de cellules, non neuronales, dans une région très limitée du cerveau. Il n'est pas exclu d'imaginer que d'autres modifications de comportement puissent être liées de façon globale à des changements intervenus dans l'organisation ou le fonctionnement de cellules gliales.
Dans le système nerveux central, les cellules gliales sont soit des cellules d'"emballage" (astrocytes, oligodendrocytes et microglie), soit des cellules épendymaires qui recouvrent la paroi des cavités (ventricules et canal de l'épendyme).
Plus de détail sur les cellules gliales
Situé dans la boîte crânienne, le cerveau est le siège des fonctions supérieures (fonctions cognitives, sens, réponses nerveuses) et végétatives. C'est donc un organe essentiel qui assure la régulation de toutes les fonctions vitales. Le cerveau reçoit des informations provenant de l’ensemble du corps humain via des nerfs afférents, les intègre et les analyse puis répond en émettant de nouveaux signaux qui redescendent vers les parties du corps concernées par les nerfs efférents.
Le cerveau humain comporte environ 100 milliards de neurones et possède un poids moyen de 1.300 à 1.400 grammes ; à la naissance il pèse 1/4 du poids final, à 6 mois 1/2 et à 2 ans 80% (c'est le début de la période critique qui va durer jusqu'à environ 8/10 ans)
Il contient également des cellules gliales et des astrocytes. Il baigne dans le liquide céphalo-rachidien.
le télencéphale, la partie supérieure, composé du cortex cérébral et du striatum ;
le diencéphale qui contient le thalamus et l'hypothalamus (glandes endocrines) ;
le mésencéphale ;
le cervelet (coordination motrice et apprentissage des mouvements routiniers) ;
le pont de Varole ;
le bulbe rachidien (contrôle des mouvements réflexes et végétatifs : respiration, modération du rythme cardiaque, régulation des muscles lisses artériels) qui communique avec la moelle épinière.
le lobe frontal (motricité, mémoire, raisonnement) ;
le lobe pariétal (toucher) ;
le lobe temporal (ouïe, odorat) ;
le lobe occipital (vision) ;
le lobe limbique
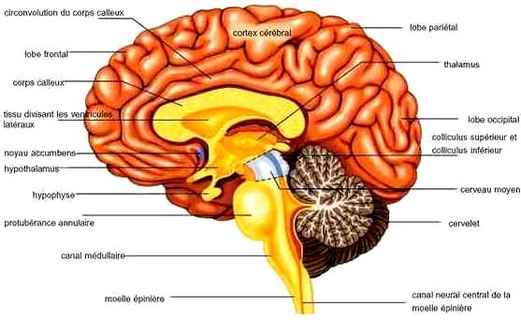
Lorsque l'on dissèque le cerveau, on remarque que ce dernier est principalement constitué d'une matière blanchâtre à laquelle on donne le nom de substance blanche. À la surface du cerveau se trouve au contraire une fine pellicule de matière grisâtre : c'est le cortex cérébral, ou substance grise.
1 - par le crane ;
2 - le liquide céphalo-rachidien qui entoure tout le cerveau et va jusqu'à la moëlle épinière (d'ou les ponctions lombaires); ce liquide contient des substances qui ne passeront jamais dans le sang.
3 - (très difficile à voir à l'oeil nu mais très important)La barrière hémato-encéphalique. (reprendre à 16')
Chez l'homme, les deux hémisphères cérébraux sont particulièrement développés. Ils ressemblent à une étoffe chiffonnée, ramassée sur elle-même, d'où le fait qu'on aperçoive une succession de bosses et de creux : les gyri et les sulci.
La vie est apparue il y a environ 4 milliards d'années et les premiers cerveaux il y a environ 400 millions d'années.
Un réseau complexe : Le cerveau ressemble à un immense réseau de fils électriques. Tous ces fils sont en fait les "queues" des neurones, l'élément de base du cerveau. Notre cerveau en contient plus de 100 milliards, dont chacun communique avec 10 000 autres de ses voisins.
1 - Plaque neurale
2 - Gouttière neurale
3 - Tube neural ou tube nerveux
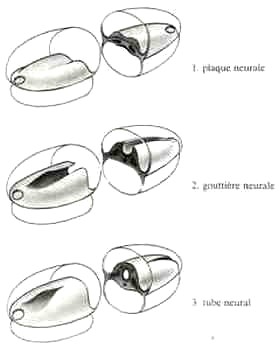
On dit souvent que le cerveau est une représentation du monde — imago mundi — et que, réciproquement, le cerveau agit le monde — anima mundi — selon les programmes innés ou acquis. La stabilité de ces images et de ces programmes exige un isolement auquel contribue la barrière hémato-encéphalique.
Neurologue, neurobiologiste, neurochimiste, neurophysiologiste, neuroendocrinologiste, neurochirurgien, neuropsychiatre, neuropédiatre, neuropsychologue, et depuis quelques temps, neuroplasticien.
a) - des activités musculaires du mouvement volontaire global
b) - des activités musculaires toniques de la posture
c) - des activités musculaires réflexes du maintien de l’équilibre.
Il est placé dans la fosse crânienne postérieure ou fosse cérébelleuse de la boite crânienne. Il est disposé sous une forte toile fibreuse dépendant de la dure-mère et appelée la tente du cervelet. Pour lui-même il est recouvert par les 3 méninges classiques. Il est entouré par l’espace sous-arachnoïdien contenant le liquide céphalo-rachidien. Autour du cervelet, cet espace constitue des cavités plus vastes appelées : citernes. Dans cet espace étroit et rigide, les pathologies expansives (vasculaires, infectieuses et surtout, tumorales), ont un retentissement rapide sur le tronc cérébral.
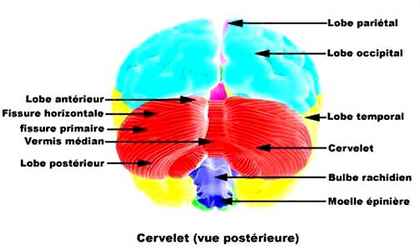
Plus de détail sur le cervelet
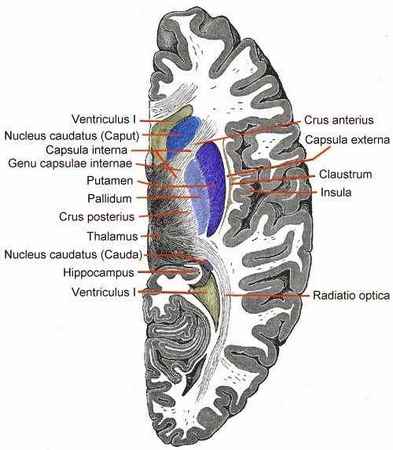
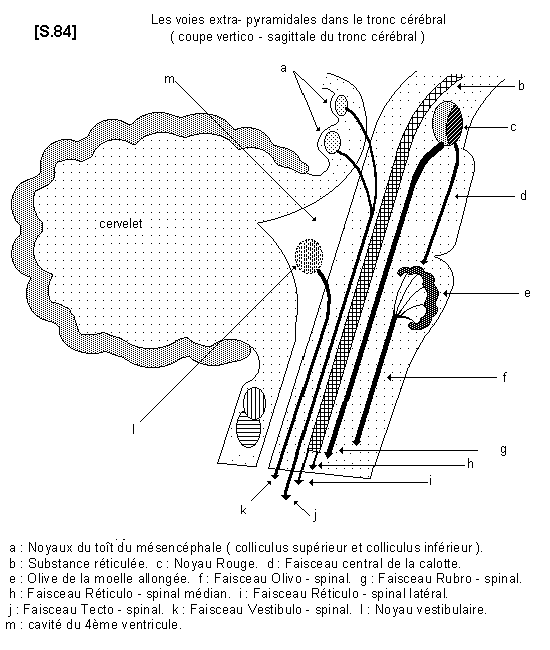
Les collicules supérieurs font partie du tectum, région qui est étroitement interconnectée avec les noyaux du gris périaqueducal et indirectement avec le nucleus tractus solitarius et le noyau parabrachial. L'implication du collicule supérieur dans le comportement lié à la vision est bien connue. Cependant, à l'exception notable des travaux de Bernard Strehler, Jaak Panksepp et Bjorn Merker, il est rare qu'on considère que ces structures peuvent jouer un rôle dans le processus de l'esprit et du soi". L'anatomie du collicule supérieur est fascinante et nous invite à conjecturer ce que cette structure est supposée accomplir. Il a sept couches : les couches I à III sont dites « superficielles » ; les IV à VII « profondes ». Toutes les connexions arrivant aux couches superficielles et en partant ont à voir avec la vision. La principale couche superficielle, la II, reçoit des signaux issus de la rétine et du cortex visuel primaire. Ces couches assemblent une carte rétinotopique du champ visuel du côté opposé.
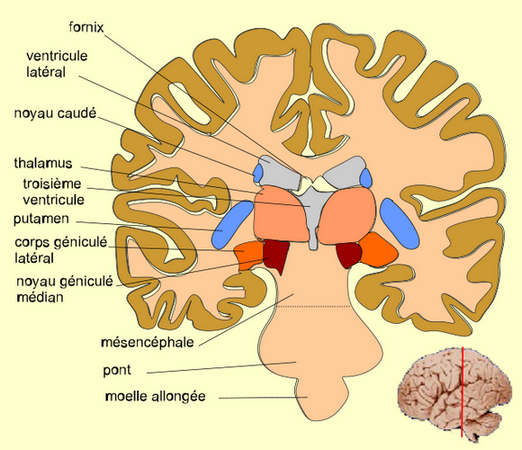
Le lobe frontal responsable du comportement d'un individu et contenant par ailleurs l'air motrice primaire qui commande tous les mouvements volontaires (située au niveau de la circonvolution frontale ascendante). Il s'agit d'une aire motrice qui est à l'origine du faisceau pyramidal permettant la motricité volontaire.
Le lobe pariétal siège du cortex pariétal situé en haut de chaque hémisphère, sur le côté et intervenant dans la connaissance du corps. Il permet à un individu d'avoir des repères spatiaux et de contrôler ces gestes et ses mouvements. Ainsi, l'apraxie est un trouble acquis de l'exécution de mouvements, troubles faisant suite à une lésion localisée, entre autres au niveau du lobe temporal. En réalité les apraxies surtout quand elles sont idéatoires font suite à une atteinte du lobe temporal et du lobe pariétal. Le lobe pariétal comprend essentiellement l'air sensitif c'est-à-dire la partie du cortex cérébral qui assure la réception des informations en provenance de la périphérie du corps et en particulier de la peau.
Le lobe temporal situé sur le côté de l'hémisphère, en dessous du cortex pariétal et qui participe aux différentes fonctions du cerveau à savoir le goût, l'audition, l'olfaction, la mémoire, le langage et la vie végétative autrement dit le fonctionnement automatique des viscères.
Le lobe occipital qui se trouve en arrière de l'hémisphère et qui contient l'aire visuelle (aire calcarine ou aire visuelle située à la face interne du lobe occipital) recevant et analysant les informations provenant du globe oculaire et plus précisément de la rétine.
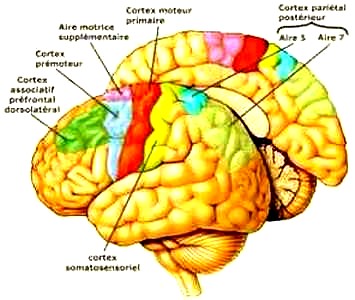
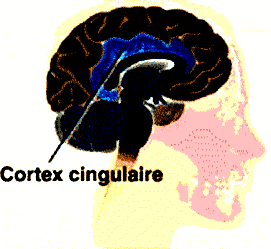
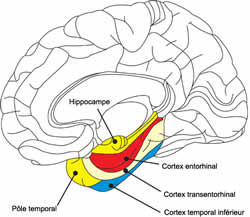
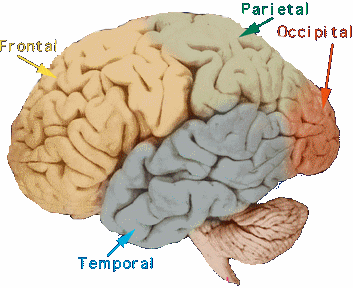
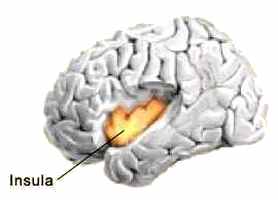
Au niveau du cortex cérébral, la principale région impliquée dans les sentiments est le cortex insulaire, qui représente une partie assez volumineuse mais bien cachée du cortex cérébral, située sous les opercules frontal et pariétal. L'insula – qui, comme son nom l'indique, ressemble à une île – comporte plusieurs gyrus. Sa partie avant est ancienne, est liée au goût et à l'odorat, et, comme pour compliquer encore un peu plus les choses, est une plateforme non seulement pour les sentiments, mais aussi pour le déclenchement de certaines émotions. Elle sert de déclencheur à une émotion des plus importantes : le dégoût. C'est l'une des plus anciennes du répertoire des émotions. Elle a commencé comme moyen automatique de rejeter de la nourriture potentiellement toxique et de l'empêcher de pénétrer le corps. Les êtres humains peuvent être dégoûtés non seulement par de la nourriture avariée et la mauvaise odeur qui l'accompagne, mais aussi par diverses situations dans lesquelles la pureté des objets ou le comportement sont compromis et où il y a « contamination ». Ils sont aussi dégoûtés par la perception d'actions moralement répréhensibles. Par suite, de nombreuses actions figurant dans le programme humain du dégoût, dont ses expressions du visage typiques, ont été cooptées par une émotion sociale : le mépris. C'est souvent une métaphore du dégoût moral.
L'insula est une partie du cortex cérébral dont la position en repli à l’intérieur des circonvolutions cérébrales la rend moins accessible. Voilà pourquoi elle est restée méconnue pendant longtemps, jusqu’à ce que des neurobiologistes comme Antonio Damasio mettent en évidence son rôle dans nombre de nos sentiments. Très branchée sur nos réactions viscérales, elle était parfaitement positionnée pour nous faire prendre consciences de ces bouleversements corporels internes associés à la moindre de nos émotions.

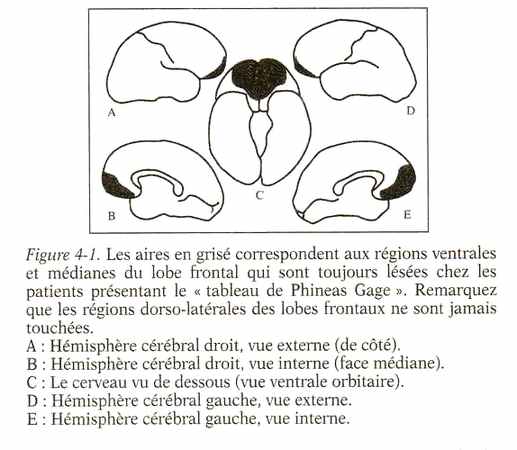
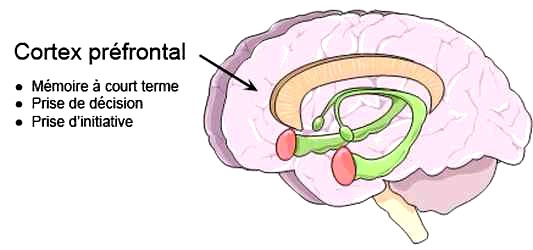
Il joue un rôle essentiel dans la mémoire épisodique.
Le LTM n'est pas un groupe de rap, mais un ensemble d'environ un milliard de neurones logés au creux du lobe temporal, là où vient se glisser l'empreinte digitale du pouce quand vous fermez votre poing, pour reprendre l'image du gant de boxe. Les patients privés de cette structure sont amnésiques et éprouvent de grandes difficultés à imaginer des situations différentes de celle de l'instant présent : « Que feriez-vous si vous vous retrouviez au milieu de l'autoroute ? — Je ne sais pas". » Le LTM n'est pas une structure cérébrale homogène, mais un assemblage de petites régions organisé de façon hiérarchique avec, à sa tête, l'hippocampe et, plus bas, le cortex entorhinal, le cortex périrhinal et le gyrus parahippocampique. L'hippocampe s'enroule sur lui-même comme l'animal marin du même nom et reçoit ses informations du cortex entorhinal, qui lui-même reçoit ses informations du cortex périrhinal et du gyrus parahippocampique, et, dans une moindre mesure, d'autres régions de haut niveau de la voie visuelle ventrale.

les androgènes, impliquées dans le développement des caractères sexuels;
les minéralocorticoïdes (aldostérone, corticostérone, désoxycortisone) qui régulent l'équilibre osmotique du corps;
les glucocorticoïdes (cortisone, hydrocortisone, prednisone) qui, outre leur activité anti-inflammatoire et immunosuppressive, stimulent la synthèse du glucose et augmentent la mobilisation des acides gras et des protéines pour répondre à la demande métabolique plus élevée engendrée par un stress.
Le rôle des neurotransmetteurs aminergiques - ou monoamines - dans le mécanisme de la dépression fut examiné dès les années 60. En effet, depuis la constatation de l’efficacité des antidépresseurs tricycliques (inhibiteurs de la recapture de la sérotonine et de la noradrénaline) dans la dépression, l’hypothèse d’un dysfonctionnement monoaminergique central - en particulier sérotoninergique – a longtemps prédominé. Plus récemment, des données nouvelles issues de la neuro-imagerie, de la neurophysiologie et de la biologie cellulaire et moléculaire ont permis d’associer la dépression à différents phénomènes neurobiologiques notamment un dysfonctionnement de la neuroplasticité.
La neuroplasticité correspond à des changements d’organisation et de structure de certains éléments neuronaux, produisant une modification ou une modulation de leur fonction (neurogenèse, apoptose, plasticité synaptique, réorganisation de la composition de réseaux neuronaux…). Les modifications seraient fonction de facteurs de l’environnement dans lequel vit le sujet (ex : stress, alcool) et de l’environnement dans lequel se trouve le système nerveux (ex : taux de glucocorticoïdes).
Sachant que la sérotonine (5-HT) régule plusieurs aspects de la plasticité cérébrale et que la plupart des antidépresseurs efficaces facilitent la transmission 5-HT, il a été proposé que ces psychotropes exercent leur effet thérapeutique en favorisant les processus neuroplastiques.
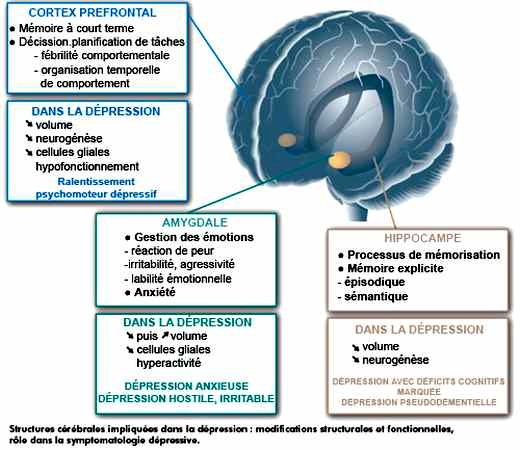
Une des régions du cortex préfrontal les plus affectées à la fois dans la dépression et la phase maniaque (d’un maniaco-dépressif) est le cortex ventromédian (aussi appelée « subgenual » parce que située sous le genou du corps calleux). Cette région située en profondeur des lobes frontaux, de chaque côté de la ligne centrale séparant les deux hémisphères, nous permet de passer d’un affect à un autre tout en étant très impliquée dans le sentiment de plaisir et de renforcement positif.
On sait en effet que ces structures sont toutes inter-reliées par des voies nerveuses. L’amygdale, pouvant générer nombre d’émotions reliées à la peur, ferait surgir un sentiment négatif, les lobes préfrontaux réanimeraient des mauvais souvenirs correspondants à un tel sentiment, le cortex cingulaire antérieur nous empêcherait de penser à autre chose et le thalamus favoriserait l’activité des circuits qui formeraient cette «boucle de la dépression…
Des chercheurs américains viennent de mettre en évidence chez la souris "le circuit" de l’anhédonie, cette difficulté à ressentir des émotions positives et expérimenter le plaisir. Cette découverte laisse envisager la possibilité de nouveaux traitements pour certaines dépressions sévères.
C’est en cherchant à mieux comprendre les circuits cérébraux impliqués dans l’apparition de symptômes dépressifs induits par un stress chronique que Rob Malenka et son équipe ont fait cette découverte. En effet, le stress chronique induit la production de mélanocortines, hormones impliquées dans la pigmentation de la peau mais aussi dans la modulation de la prise alimentaire. Or, il existe de nombreux récepteurs de mélanocortine dans le noyau accumbens, une région du cerveau impliquée dans le "mécanisme de récompense". Quand ce circuit fonctionne normalement, il permet de ressentir du plaisir suite à l’accomplissement d’un objectif ou d’expériences agréables (liées à la nourriture, au sexe…). A l’inverse, quand ce circuit est défectueux, pas de plaisir. On parle alors d'anhédonie.
Mais jusqu’à présent, on ne savait pas si la modulation de l’activité de mélanocortine pouvait influer sur ce circuit de récompense et donc sur la "joie de vivre".
L'exemple type du « traitement profond » est un traitement sémantique, qui traite du sens des items à mémoriser. Or les sujets âgés ont du mal à utiliser spontanément des stratégies de traitement sémantique au moment de l'encodage. Ceci a été démontré dans des études qui demandent à des sujets de mémoriser des séries de mots appartenant à différentes catégories sémantiques, les mots étant présentés en désordre (par exemple, « jasmin », « cartable », « voiture », « tulipe », « train », « cahier », « avion », « rose »...). Les sujets jeunes regroupent les mots par catégorie sémantique dès le premier rappel, tandis que les sujets âgés le font moins spontanément.
Les tests de reconnaissance avec distracteurs sémantiques (par exemple, « dahlia » ou « lilas » pour la catégorie des fleurs) entraînent plus de fausses reconnaissances chez les sujets âgés que chez les jeunes, ce qui pourrait provenir d'un encodage insuffisamment précis et distinctif. L'encodage chez les sujets âgés se ferait à un niveau relativement global, qui leur permettrait de rejeter des distracteurs non liés sémantiquement (par exemple, « chameau »), alors que la possibilité de discriminer
correctement des items proches sémantiquement exige un encodage plus élaboré et plus coûteux en ressources attentionnelles.
Des aides fournies par l'expérimentateur lors de l'encodage permettent de réduire, voire d'éliminer, les différences liées à l'âge. La compensation est d'autant plus efficace que la consigne demande explicitement un traitement sémantique (par exemple, faire une petite phrase avec le mot ou donner un synonyme).
L'effet délétère de l'âge sur la récupération d'informations encodées après un traitement profond persiste chez les sujets les plus âgés, comme le montrent les études ayant inclus un groupe de sujets d'âge intermédiaire (une cinquantaine d'années) en plus des sujets jeunes et des sujets âgés classiquement comparés. En d'autres termes, les sujets d'âge intermédiaire profitent des aides et obtiennent des performances équivalentes à celles des jeunes, alors que les plus âgés améliorent leurs performances mais restent moins « bons » que les autres sujets. Par contraste, le rappel d'informations traitées de façon superficielle lors de l'encodage (par exemple, en comptant le nombre de voyelles dans les mots) semble diminuer dès l'âge intermédiaire.
Le précurseur est empaqueté dans des granules grâce à l'intervention de l'appareil de Golgi. Ces granules sont transportés vers les terminaisons nerveuses par le flux axonal. Au cours du transport, le précurseur subit une maturation qui conduit à l'apparition de l'hormone sous sa forme définitive. La libération de l'hormone se fait par accolement et rupture du granule à l'extérieur de la membrane cellulaire (exocytose). Il ne faut pas confondre le transport axonal du granule avec la conduction de l'influx nerveux. Ce dernier, parvenu à la terminaison sous la forme d'un potentiel d'action, provoque à ce niveau une entrée de calcium dans la cellule. Cet afflux de calcium est le principal responsable de l'exocytose.
1 - Lorsque nous exerçons une activité qui fait réagir ensemble des neurones spécifiques, ceux-ci libèrent un médiateur chimique, le FNIC, qui raffermit les connexions entre ces neurones et garantit la simultanéité de leurs réactions.
2 - Le FNIC favorise aussi la croissance de la mince couche graisseuse qui entoure les neurones et accélère la transmission des signaux électriques.
3 - Au cours de la période critique, le FNIC excite le nucleus basalis ou noyau basal de Meynert (NBM), la partie du cerveau qui nous permet de fixer notre attention, et le maintient en état de vigilance pendant toute la durée de cette période. L'activité du noyau basal nous aide également à mémoriser les événements que nous vivons. Elle permet à la diversification des aires et aux changements afférents d'avoir lieu sans effort.
— C'est comme un professeur dans le cerveau qui nous dirait : « Voici ce qui est important, ce qu'il faut que tu retiennes pour l'examen de la vie », explique Merzenich.
Il a baptisé le noyau basal et le système de maintien de l'attention « système de contrôle modulateur de la plasticité ». Lorsqu'il est activé, ce système met le cerveau en état d'extrême plasticité.
4 - Le quatrième et dernier service que rend le FNIC, c'est d'aider à mettre fin à la période critique. Une fois que la consolidation des connexions essentielles est achevée, il y a dans le cerveau un besoin de stabilité, et donc de baisse de la plasticité. Quand la substance chimique du FNIC est libérée en quantité suffisante, cela interrompt l'activité du noyau basal et clôt cette période faste d'apprentissage sans effort. Dès lors, le noyau basal n'agira plus que s'il nous arrive quelque chose d'important, de surprenant, d'inédit, ou si nous fournissons un gros effort d'attention.
Les travaux de Merzenich sur la période critique et le FNIC l'ont aidé à bâtir une théorie susceptible d'expliquer la multiplicité des problèmes rencontrés par les jeunes enfants génétiquement prédisposés à l'autisme. Durant la période critique, certaines situations surexcitent leurs neurones. Le médiateur chimique du FNIC est alors libéré massivement avant tenue. Toutes les connexions, et non les plus importantes, s'en trouvent renforcées, ce qui met fin prématurément à la période critique. Les connexions sont scellées en l'état, et le cerveau de l'enfant se retrouve doté d'aires cérébrales non sélectives. C'est ce qui explique les troubles de la croissance qu'on observe chez ces malades. Leur cerveau hypersensible est en état d'hyperexcitation permanente. S'ils entendent une seule fréquence sonore, tout leur cortex auditif réagit". C'est sans doute ce qui se passait quand les oreilles « bioniques » de Lauralee percevaient certaines musiques. D'autres jeunes autistes présentent une hypersensibilité tactile qui les tourmente, par exemple lorsque l'étiquette de leurs vêtements est directement en contact avec la peau. La théorie de Merzenich explique aussi la fréquence de l'épilepsie chez les autistes. Le renforcement indifférencié de connexions neuronales en surnombre conduit à l'« embrasement » du cerveau à la moindre sollicitation. Cette théorie permet aussi de comprendre pourquoi ces enfants ont un cerveau plus gros". La libération excessive du médiateur chimique accroît considérablement la couche graisseuse qui enveloppe leurs neurones.
À partir de ce constat, il restait à Merzenich à comprendre ce qui provoque cette surexcitation des jeunes neurones et le dérèglement chimique qui en résulte.
Différentes études lui faisaient penser qu'un facteur environnemental était en cause. L'une d'elles montrait qu'à Francfort, en Allemagne, plus les enfants vivent à proximité du bruyant aéroport, moins ils sont intelligents. Une étude similaire montrait des résultats comparables avec des enfants vivant dans les parages d'une autoroute, aux environs de Chicago. Merzenich commença à s'interroger sur le rôle que pouvait jouer cette pollution sonore, phénomène historiquement nouveau, sur des enfants prédisposés à l'autisme. Le bruit de fond continu d'origine industrielle est parfois appelé « bruit blanc ». Il se compose de fréquences multiples qui ont un fort impact sur le cortex auditif.
— Les jeunes enfants grandissent dans un environnement de plus en plus bruyant, dit-il. Le vacarme est permanent. Il provient des ventilateurs intégrés dans l'électronique, des climatiseurs, du chauffage et bien sûr de la circulation automobile. Comment le cerveau ne serait-il pas affecté ?
Pour valider cette hypothèse, son groupe a exposé de jeunes rats à des émissions de bruit blanc tout au long de la période critique. Le cortex de ces animaux s'en est trouvé dévasté.
— À chaque pulsation sonore, commente Merzenich, on assiste à une excitation générale du cortex. Pas un neurone n'est épargné.
Les décharges électriques simultanées de neurones si nombreux entraînent une suractivité au niveau du FNIC, et, conséquemment, la fin prématurée de la période critique, comme le prévoyait le modèle. Au terme de l'expérience, les jeunes rongeurs présentent des aires auditives uniformisées et des neurones non différenciés qui réagissent à n'importe quelle fréquence.
Merzenich s'est aperçu que ces jeunes rats, comme les enfants autistes, étaient fréquemment sujets à l'épilepsie et qu'ils avaient des crises à l'audition de la parole normale. (Chez l'homme, ce sont les lumières stroboscopiques desh concerts rock qui déclenchent les crises. Ces émissions d'éclairs lumineux constituent une lumière blanche qui comporte elle aussi de multiples fréquences.) Merzenich disposait maintenant d'un modèle animal pour l'autisme.
Des explorations scanographiques récentes ont confirmé que le traitement cérébral des données auditives était effectivement anormal chez les jeunes autistes. Selon Merzenich, un enfant dont le cortex est indifférencié éprouve d'énormes difficultés à fixer son attention. Quand on lui demande de se concentrer sur quelque chose, les informations qui lui parviennent se télescopent, il est en proie à une grande confusion. C'est la raison pour laquelle les jeunes autistes se retirent du monde et se réfugient derrière une cuirasse. Sous une forme atténuée, le même problème se manifeste chez les personnes qui présentent des troubles de l'apprentissage.
Voir aussi : dossier autisme
Les noyaux de cette région qui se placent à côté des noyaux du raphé sont constitués de cellules de grande taille (gigantocellulaires) à nombreuses arborisations.
1. le noyau bulbaire caudal (central) ;
2. le noyau gigantocellulaire (bulbaire) ;
3. le noyau réticulaire pontin ou pontique (protubérantiel) divisé en :
- noyau caudal de la réticulée pontique,
- noyau oral de la réticulée pontique ;
4. le noyau cunéiforme (mésencéphalique) :
- le noyau principal, interne, (de Burdach) est impliqué dans le traitement de la sensibilité ( infos) ;
- le noyau accessoire ou sous-cunéiforme, externe, se connecte avec le cervelet.
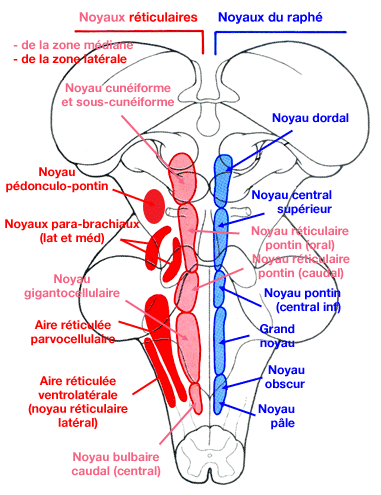
Pour simplifier, la région latérale (qui s'arrête avant le mésencéphale) reçoit des projections nerveuses de toutes sortes, les filtre et les transmet aux régions médiales qui activent ou inhibent le système nerveux.
1. le noyau parvocellulaire (bulbe et pont) ;
2. le noyau parabrachial (protubérance et mésencéphale) ou aire parabrachiale.
1. le noyau réticulaire latéral - noyau latéral du bulbe - (bulbaire) ;
2. le noyau réticulaire paramédian - paramédian de la réticulée ou central du bulbe - (bulbaire) ;
3. le noyau réticulo-tegmental pontin ;
4. le noyau pédonculo-pontin (pédiculo-pontin ou pédonculaire).
En effet, l'hippocampe proprement dit est fait des régions pourvues de neurones pyramidaux étroitement groupées, principalement les aires CA1, CA2 et CA3 (CA pour Corne d'Ammon, à cause de sa ressemblance avec la forme des cornes de bélier du dieu Ammon de la mythologie). C'est ce que l'on appelle le circuit ou la boucle trisynaptique de l'hippocampe.
L'entrée de l'information dans cette boucle unidirectionnelle se fait par les axones du cortex entorhinal nommés fibres perforantes (ou voie perforante, car elle passe au travers du subiculum et de l'espace qui le sépare du gyrus dentelé). Ces axones font la première connexion de la boucle avec les neurones du gyrus dentelé.
Des cellules granuleuses du gyrus dentelé partent les fibres moussues qui font la deuxième connexion de la boucle avec les dendrites des cellules pyramidales de l'aire CA3.
Les axones des cellules de l'aire CA3 se ramifient en deux : une partie forme les fibres commissurales se dirigeant vers l'hippocampe controlatéral via le corps calleux et l'autre partie forme les collatérales de Schaffer qui font la troisième connexion de la boucle avec les cellules de l'aire CA1.
C'est dans ces synapses que la mémoire spatiale associée à l'hippocampe semble être encodée. Cette région démontre aussi une forte propension à la potentialisation à long terme (PLT), bien que l'on observe ce phénomène à plusieurs autres endroits dans l'hippocampe ainsi que dans le cortex.
Finalement, les axones des cellules de CA1 se dirigent vers les neurones du subiculum et les neurones du cortex entorhinal. La partie réceptrice de la formation hippocampique est donc constituée par le gyrus denté et sa partie émettrice par le subiculum. Les axones des grands neurones pyramidaux du subiculum se dirigent ensuite vers les noyaux sous-corticaux par la fimbria, mince lame de substance blanche au bord interne de l'hippocampe. Finalement, l'information retournera vers les aires corticales sensorielles d'où elle était partie avant d'être traitée par l'hippocampe.
Dans les années 1970, une découverte étonnante fit la démonstration que l'hippocampe du rat contient une véritable carte spatiale de l'environnement où il évolue. Certains neurones pyramidaux de la région CA1 vont en effet entrer en activité uniquement lorsque le rat va se trouver dans une région précise de son environnement.
Ces cellules, dites "cellules de lieu", sont au nombre de 1,000,000 dans l'aire CA1 du rat, de sorte que si chacun se voit assigné un endroit précis dans l'espace, ils peuvent former une carte cognitive très précise capable d'informer l'animal à tout moment de l'endroit où il se trouve. De plus, l'exploration d'un nouvel environnement va induire la formation d'une nouvelle carte cognitive qui peut être très stable (semaines, mois).
Une des fonctions possibles de ces représentations spatiales dans l’hippocampe serait, selon O’Keefe et Nadel qui les ont mis en évidence, de créer un contexte sur lequel peuvent se greffer les souvenirs. Un événement mémorisé peut alors devenir autobiographique, c’est-à-dire situé dans le temps et dans l’espace, ce qui expliquerait le rôle fondamental de l’hippocampe dans la mémoire épisodique chez l’humain
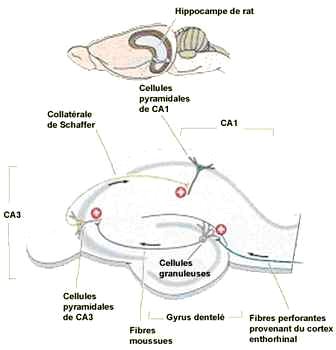
Par exemple, lors d'un souper bien arrosé avec des amis, le souvenir des visages des personnes présentes, du goût du vin ou de la musique qui jouait est distribué dans les différentes aires visuelles, olfactives et auditives du cerveau, mais sont liés ensemble par l'hippocampe pour former un " épisode " au lieu de demeurer une collection de souvenirs séparés.
L'hippocampe joue donc un rôle primordial dans la mémoire épisodique, celle qui nous permettra des années plus tard de nous rappeler cette soirée particulièrement agréable. En effet, c'est l'hippocampe qui semble nous permettre de " rejouer la scène " en réactivant ce pattern particulier d'activité de différentes régions corticales. Ce phénomène serait très important durant les rêves, ce qui expliquerait l'incorporation d'événements des derniers jours dans ceux-ci.
Mais au bout d'un certain temps, ces différentes régions corticales activées lors d'un événement deviendraient fortement liées entre elles et pourraient se passer du travail de l'hippocampe comme agent de liaison. Grâce à ce couplage, le seul souvenir d'une pièce musicale qui jouait ce soir-là pourrait nous rappeler toute la scène du souper, chacun de ces éléments pouvant servir d'indice pour faire resurgir à la conscience tout le reste.
Les souvenirs encodés depuis longtemps dans la mémoire à long terme peuvent donc se passer de l'hippocampe. C'est en particulier le cas des connaissances générales de la mémoire sémantique qui activent plutôt le cortex frontal et le cortex temporal. L'activité du lobe temporal correspondrait à l'activation du fait en question et celle du cortex frontal à son accession à la conscience.
Contrairement aux faits et aux événements cependant, notre mémoire spatiale demeurerait, elle, confinée à l'hippocampe. Et plus précisément à l'hippocampe droit. Celui-ci aurait la capacité de recréer une carte mentale de l'espace, grâce à certaines cellules dites ''cellules de lieu''
Certains souvenirs personnels très intenses, mettant en jeu ce qu'on appelle parfois la mémoire émotive, impliqueraient en plus de l'hippocampe une autre structure du système limbique. Il s'agit de l'amygdale, une région déjà reconnue pour gérer nos réactions de peur. D'ailleurs, plusieurs autres structures du système limbique contribuent à encoder nos souvenirs de façon durable.
Enfin, la mémoire procédurale, celle du " savoir faire " comme aller à bicyclette par exemple, ne solliciterait pas du tout l'hippocampe. Elle serait plutôt associée à des modifications dans le cervelet, les ganglions de la base et le cortex moteur, des régions justement impliquées dans le contrôle de la motricité. D'ailleurs, la mémoire procédurale n'est pas touchée par l'amnésie liée aux lésions de l'hippocampe mais elle est affectée par des dommages au cervelet et des maladies neurodégénératives comme celle de Huntington qui altèrent les ganglions de la base.
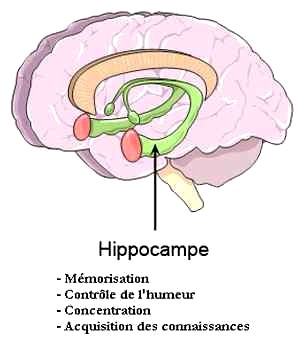
Le gyrus denté (gyrus est le terme médical signifiant circonvolution) est une région hippocampique particulièrement intéressante dans les troubles thymiques. En effet, il s’agit d’une des rares zones susceptibles de synthétiser de nouveaux neurones à l’âge adulte. Or cette neurogenèse est perturbée lors d’un épisode dépressif et certains traitements antidépresseurs sont susceptibles de la restaurer. Ces données récentes donnent à l’hippocampe un rôle central dans la physiopathologie des troubles thymiques et de leur traitement.
Le gyrus denté est bordé, en haut, par une lame de substance blanche, appelée fimbria de l’hippocampe.
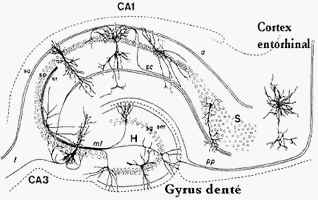
À la précision des circuits nerveux de notre cerveau câblé, on peut opposer la soupe diffuse de notre cerveau hormonal. Mais cette opposition n'est que théorique puisque dans la " vraie vie ", les deux se complètent admirablement bien.
Certaines substances de nature le plus souvent peptidique, lorsqu'elles sont injectées en quantité minime dans des régions particulières du cerveau de l'animal, déclenchent une série d'actes moteurs qui constituent ce que l'on appelle un comportement : manger, boire, faire l'amour, etc. Nous en donnons ici quelques exemples.
Aujourd'hui, grâce aux techniques génétiques modernes, il est possible de synthétiser des hormones de croissance. Ce traitement désormais est considéré comme sûr, même s'il comporte certains risques, comme tout traitement. Il pose en outre certains problèmes éthiques, l'hormone de croissance pouvant être utilisée pour augmenter des performances athlétiques ou pour grandir artificiellement au-delà de la taille normale.
L'hypothalamus est aussi influencé par le stress. Ce dernier peut provenir de l'intérieur ou de l'extérieur du corps. Les facteurs externes de stress peuvent être, par exemple, le hurlement d'une radio, une piqûre d'épingle ou la vue d'une balle de tennis qui vous arrive droit dessus. Le stress interne peut être provoqué par des sentiments dépressifs ou de l'anxiété. Quand le stress intervient, l'hypothalamus envoie un message à la glande pituitaire(ou hypophyse), ce qui active la glande surrénale et augmente la production d'adrénaline, qui, à son tour, provoque l'augmentation du rythme cardiaque, la dilatation des pupilles (pour une meilleure vision), la constriction des vaisseaux sanguins cutanés (afin que plus de sang afflue vers les muscles et le cerveau) et, enfin, l'interruption de la digestion.
Tout cela marche parce que la corticolibérine (corticotropin releasing factor ou CRF en anglais) produite par l'hypothalamus stimule la glande pituitaire, qui produit de l'adrénocorticotrophine (adrenocorticotropic hormone ou ACTH en anglais), laquelle, à son tour, stimule la glande surrénale qui produit l'adrénaline.
Comme pour les autres parties du cerveau, les activités de l'hypothalamus se déroulent sans que vous en soyez conscient. L'hypothalamus vous fait transpirer quand il fait chaud pour vous rafraîchir et vous fait frissonner quand il fait froid pour augmenter la production de chaleur. C'est votre hypothalamus qui vous indique si vous avez faim.
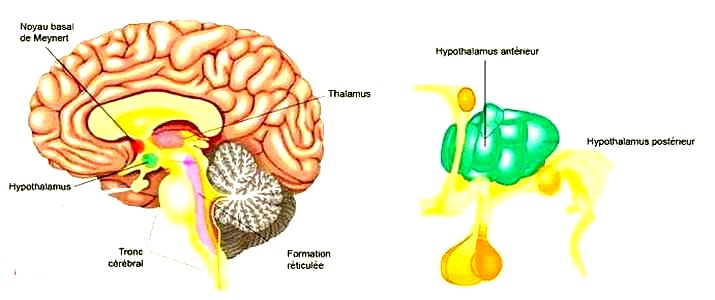
Axe Hypothalamo-hypophysaire
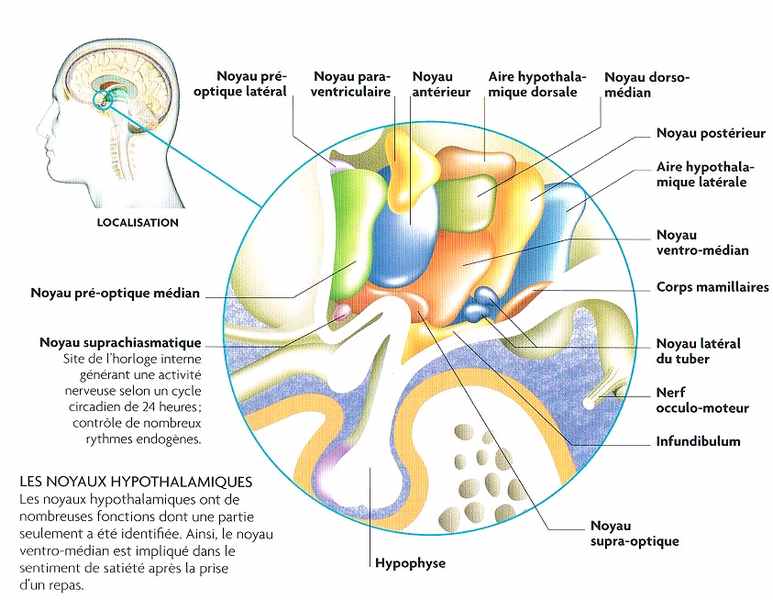
En dehors de cas cliniques exceptionnels, aucune technique ne permet de mesurer l'activité du cerveau humain à l'échelle de la milliseconde et du millimètre de façon non invasive, c'est-à-dire sans chirurgie. Par contre, une simple électrode scotchée à la surface de la tête suffit à mesurer toutes les millisecondes le signal électrique généré par les neurones. C'est la technique de l'électroencéphalographie inventée par Hans Berger — la fameuse EEG. Ce signal est dû aux mouvements des ions au voisinage des canaux ioniques qui entraînent des accumulations de charges négatives et positives à différents endroits le long du neurone. L'existence simultanée et à faible distance l'un de l'autre d'un excès de charges positives et de charges négatives crée alors ce que l'on appelle en physique un dipôle, qui modifie le potentiel électrique autour du neurone. Lorsque ce phénomène se produit en même temps dans des millions de cellules nerveuses situées côte à côte, la modification du potentiel devient suffisamment forte pour qu'elle puisse être détectée depuis la surface du scalp, à quelques centimètres de distance. Mais le signal mesuré par l'électrode correspond à l'activité électrique moyenne d'une population de neurones très vaste, distribuée sur plusieurs centimètres carrés de cortex. Il s'agit donc d'une mesure globale, qui ne renseigne pas sur ce que fait chaque neurone en particulier. C'est un peu comme le brouhaha que vous entendez quand vous passez aux abords d'un stade un soir de match de foot. Vous n'entendez pas les paroles prononcées par chacun des spectateurs, mais plutôt une sorte de clameur qui suffit tout de même à savoir quand un but est marqué et par quelle équipe. L'EEG mesure la clameur des neurones et permet de suivre en direct l'action qui se déroule dans le cerveau.
Plus de détail sur l'imagerie cérébrale
Activités électriques cérébrales
le stress mental chronique, un état affectant de nombreux systèmes cérébraux, au niveau du néo-cortex, du système limbique et de l'hypothalamus, semble conduire à la surproduction d'une substance chimique, le peptide dérivé du gène de la calcitonine ou CGRP, au sein des terminaisons nerveuses figurant dans la peau. Par suite, ce peptide recouvre de façon excessive la surface des cellules de Langherans, qui sont apparentées au système immunitaire et ont pour fonction de capter les agents infectieux et de les présenter aux lymphocytes, de telle sorte que le système immunitaire puisse éliminer ces microbes de l'organisme. Lorsqu'elles sont complètement recouvertes de CGRP, les cellules de Langerhans sont moins fonctionnelles et ne peuvent plus jouer leur rôle de gardien. Le résultat final est que le corps devient plus vulnérable aux infections, puisqu'une porte d'entrée majeure pour les agents infectieux est désormais moins bien défendue.
Et il y a d'autres exemples d'interactions entre corps et état mental. La tristesse et l'anxiété peuvent notablement altérer la régulation des hormones sexuelles, entraînant non seulement des changements dans le domaine des pulsions sexuelles, mais aussi dans celui du cycle menstruel. Le deuil, un état mental affectant de nombreux systèmes cérébraux, conduit à une dépression du système immunitaire telle que les individus sont plus vulnérables aux infections et, en conséquence directe ou non, plus enclins à développer certains types de cancer. On peut réellement mourir d'un coeur brisé.
On peut se représenter cela comme un ensemble d'appareils d'arrosage bien étudiés, chacun d'entre eux déversant sa propre substance chimique à des systèmes particuliers et, à l'intérieur de ceux-ci, à des circuits particuliers, dotés de récepteurs spécifiques en quantité donnée. Des changements dans les quantités délivrées de l'un ou de l'autre de ces neurotransmetteurs, ou même des changements dans l'équilibre relatif de ces derniers en un site cérébral particulier, peuvent influencer rapidement et profondément l'activité corticale et donner naissance à des états mentaux du type de la dépression ou de l'exaltation ou même de l'excitation maniaque. Les processus de pensée peuvent se ralentir ou s'accélérer ; les images de rappel peuvent affluer ou au contraire disparaître ; les nouvelles combinaisons d'images peuvent être multipliées ou s'interrompre. Par suite, la capacité de se concentrer sur un contenu mental particulier peut énormément varier.
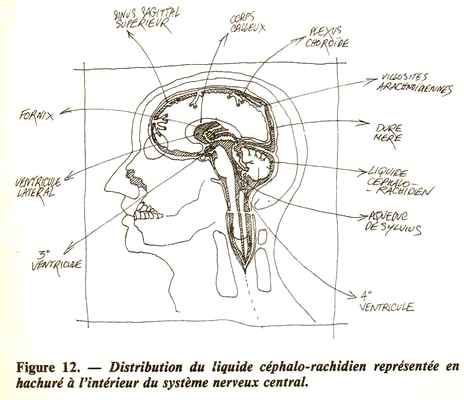
vidéo sur le liquide céphalo-rachidien
La peur et l'anxiété
L'alternance veille-sommeil
La destruction entière du locus cœruleus supprime le rêve et le sommeil paradoxal.
animation des principaux noyaux émetteurs de neurotransmetteurs