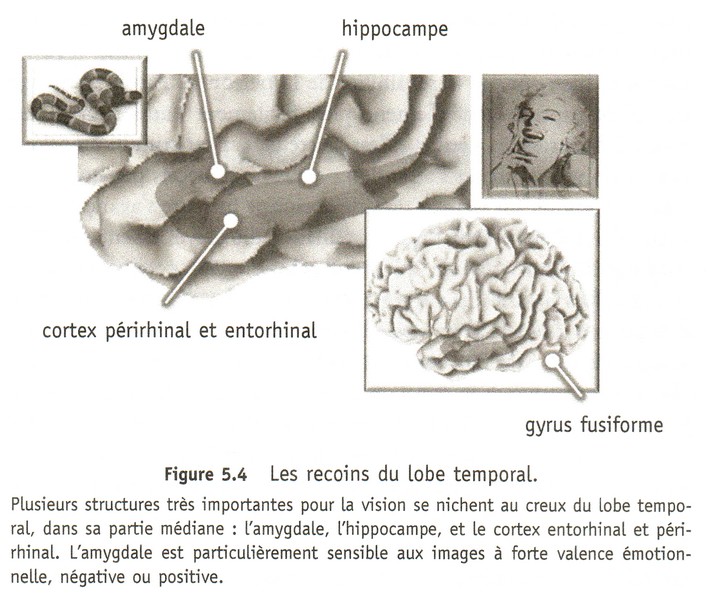
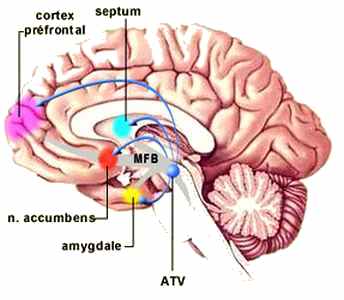
LES CENTRES DU PLAISIR: À l'arrivée d'un signal annonçant une récompense, donc après traitement sensoriel par le cortex, l'activité d’une région particulière du mésencéphale, l'aire tegmentale ventrale (ATV ; un groupe de neurones, contenant de la dopamine, situés en plein centre du cerveau), se trouve augmentée. Celle-ci libère alors de la dopamine dans le noyau accumbens, mais aussi dans le septum (le septum lucidum correspondant à la membrane fine, en position verticale, constituée de deux feuillets de forme triangulaire, qui sépare la ligne située au milieu des cornes antérieures (partie avant) des ventricules latéraux du cerveau), l'amygdale et le cortex préfrontal.
Le noyau accumbens intervient alors dans l'activation motrice de l'animal et le cortex préfrontal dans la focalisation de l'attention.
Ces régions sont reliées par ce que l’on appelle le faisceau de la récompense ou du plaisir. En terme neuro-anatomique, ce faisceau fait partie du «medial forebrain bundle (MFB)»(faisceau du cerveau antérieur médian) dont l’activation mène à la répétition de l’action gratifiante pour en consolider les traces nerveuses.
Décrit par James Olds et Peter Milner au début des années 1960, le MFB est un faisceau d’axones qui part de la formation réticulée, traverse l’aire tegmentale ventrale et continue jusqu’au noyau accumbens ainsi qu’à l’amygdale, au septum et au cortex préfrontal.
De plus, tous ces centres sont interconnectés et innervent l'hypothalamus (flèches rouges), l'informant de la présence d'une récompense. Le noyau latéral et le noyau ventromédian de l’hypothalamus sont particulièrement impliqués dans ce circuit de la récompense.
L'hypothalamus agit alors en retour non seulement sur l’aire tegmentale ventrale, mais aussi sur les fonctions végétatives et endocrines de tout le corps par l’entremise de l’hypophyse.
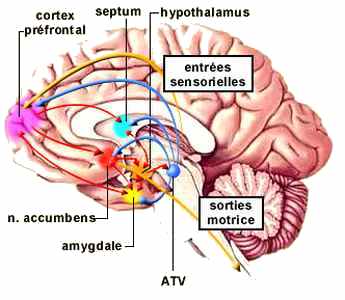
Le MFB est composé de voies ascendantes et descendantes, incluant la majorité des voies utilisant les monoamines comme neurotransmetteur. Le système dopaminergique mésocorticolimbique est l’une de ses principales composantes.
Les techniques permettant d’identifier les circuits utilisant la dopamine et de localiser leurs récepteurs ont permis de distinguer 8 voies dopaminergiques majeures dans le cerveau. Trois de celles-là sont particulièrement importantes et prennent naissance dans le mésencéphale.
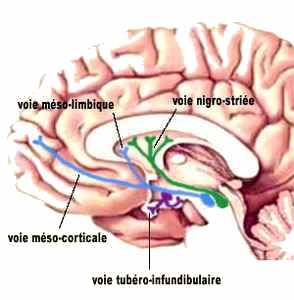
Il s’agit d’abord du faisceau dopaminergique associé au circuit de la récompense. Il est constitué par la voie méso-limbique qui part des neurones de l’aire tegmentale ventrale et innervent plusieurs structures du système limbique dont le noyau accumbens. Cette voie est importante pour la mémoire et la motivation de nos comportements. En bloquant ce faisceau, les antipsychotiques réduisent les émotions intenses provoquées par la schizophrénie, par exemple.
La voie méso-corticale part aussi de l’aire tegmentale ventrale mais innerve le cortex frontal et les structures avoisinantes. Certaines évidences montrent qu’un malfonctionnement de cette voie pourrait être à l’origine des symptômes de la schizophrénie (hallucinations, désordre de la pensée…). Le blocage de ce faisceau avec des médicaments réduit les délires psychotiques, mais aussi le fonctionnement global des lobes frontaux.
La voie nigro-striée qui projette des axones de la substance noire au striatum (noyau caudé et putamen), cette région impliquée dans le contrôle moteur. La dégénérescence de ces neurones est d’ailleurs associé aux symptômes de tremblement et de rigidité musculaire associés à la maladie de Parkinson.
Enfin, mentionnons une quatrième voie dopaminergique, la voie tubéro-infundibulaire, qui va de l’hypothalamus à l’hypophyse et influence la sécrétion de certaines hormones comme la prolactine.
2 - Et il y a le circuit de la punition. Les stimulations aversives provoquant la fuite ou la lutte activent quant à elles le circuit de la punition (ou "periventricular system" (PVS)) qui nous permet de faire face aux situations déplaisantes. Mis en évidence par De Molina et Hunsperger en 1962, ce système implique différentes structures cérébrales dont l’hypothalamus, le thalamus et la substance grise centrale entourant l’aqueduc de Sylvius. Des centres secondaires se trouvent aussi dans l’amygdale et l’hippocampe.
Ce circuit fonctionne dans le cerveau grâce à l’acétylcholine et stimule l’ACTH (« adrenal cortico-trophic hormone »), l’hormone qui stimule la glande surrénale à libérer de l’adrénaline pour prépare les organes à la fuite ou la lutte.
Il est intéressant de noter que la stimulation du circuit de la punition peut inhiber le circuit de la récompense, appuyant ainsi l’observation courante que la peur et la punition peuvent chasser bien des plaisirs.
Le MFB et le PVS sont donc deux systèmes majeurs de motivation pour l’individu. Ils incitent à l’action afin d’assouvir les pulsions instinctives et d’éviter les expériences douloureuses. Par conséquent, on peut dire que le circuit de la récompense, ainsi que celui de la punition, fournissent la motivation nécessaire à la plupart de nos comportements.
Le système inhibiteur de l'action (SIA)
Tout se gâte cependant si vous êtes un ouvrier et que le rapace à la tête du patron qui vous exploite. Vous ne pouvez fuir ou le combattre, car vous vous retrouveriez sans emploi. Alors passent les semaines et les années que vous vivez en inhibition de l'action, ce qui peut avoir des effets catastrophiques sur votre santé.
Une autre situation dans laquelle peut survenir l'inhibition de l'action est celle du déficit informationnel. En effet, pour agir efficacement, on a besoin d'un certain nombre d'informations sur le monde. Si nos apprentissages antérieurs ne nous en fournissent pas assez ou si l'on ne peut pas avoir accès à cette information, alors l'inhibition de l'action nous rattrape. C'est l'angoisse qu'éprouvent certaines personnes devant l'ordinateur par exemple, ou nous tous devant la mort.
Henri Laborit : si l’inhibition persiste, le remue-ménage biologique qu’elle entraîne, résultant en particulier de la libération de corticoïdes surrénaliens (cortisol) et de médiateurs chimiques sympathiques contractant les vaisseaux (noradrénaline), va dominer toute la pathologie: blocage du système immunitaire qui ouvrira la porte aux infections et aux évolutions tumorales, destructions protéiques à l’origine des insomnies, amaigrissement, rétention d’eau et de sels, d’où hypertension artérielle et accidents cardio-vasculaires, comportements anormaux, névroses, dépressions, etc.
Un chemin vers le plaisir - Récompense et motivation (J-P Lachaux : "Le cerveau attentif")
La dopamine semble intervenir davantage dans la motivation que dans la sensation de plaisir proprement dite. Si l'on abaisse artificiellement le niveau de dopamine chez des souris équipées du dispositif d'Olds et Milner, celles-ci continuent d'actionner spontanément la pédale, mais seulement si l'effort à fournir est minime. Le cerveau de ces souris reste donc sensible à la récompense déclenchée par l'action de la pédale, même sans dopamine. Des molécules connues pour les sensations de plaisir qu'elles procurent, comme la nicotine, l'éthanol et les cannabinoïdes présentes dans le tabac, l'alcool et Le cannabis, produisent cet effet même dans un cerveau dépourvu de dopamine. Un animal ou un être humain sans dopamine peut donc se réjouir d'une belle récompense, mais il ne fera aucun effort pour l'obtenir ; en un mot, il n'est pas motivé. Sans dopamine, je n'irai pas jusqu'au réfrigérateur pour me chercher une bière, même par 40 °C à l'ombre ; par contre, je serai ravi si vous allez m'en chercher une. C'est toute la différence entre ce que les chercheurs anglo-saxons appellent le wanting, le fait de vouloir quelque chose et d'être prêt à agir pour l'obtenir, et le liking, le fait d'apprécier cette même chose. S'il vous arrive parfois, le soir, d'être fatigué au point de ne pas parvenir à vous décoller du canapé pour alter au lit, tout en sachant pertinemment à quel point vous y seriez bien, vous avez une petite idée de ce que ressent un animal privé de dopamine.
Le circuit de la récompense, et de la motivation fait un usage immodéré d'un neurotransmetteur appelé dopamine. Un cerveau sans dopamine semble incapable de guider son action, à plus ou moins long terme, en fonction de ce qu'il aime. Grâce à des chercheurs comme Wolfram Schultz, nous commençons à savoir pourquoi.
Les neurones dopaminergiques – ceux qui utilisent la dopamine pour communiquer – ont la capacité formidable d'anticiper les récompenses : si un rat reçoit régulièrement de la nourriture dans une pièce de son enclos, le simple fait, pour cet animal, de pénétrer dans cette pièce finit par activer les neurones dopaminergiques du circuit de récompense. Au fur et à mesure que le rat apprend à connaître son environnement, ces neurones réagissent même de plus en plus tôt : d'abord au moment où l'animal rentre dans le couloir qui mène à la salle du repas, puis au moment où il aperçoit la porte de ce couloir, etc. En somme, si un stimulus, ou une situation, est systématiquement suivi d'une récompense, les neurones qui ne réagissaient au départ qu'à l'obtention de cette récompense finissent par réagir dès que la situation annonciatrice se produit. Les neurones dopaminergiques sont donc à l'affût du moindre indice indiquant une possible récompense... ou une possible punition. Au moindre signe favorable ou défavorable, ils envoient immédiatement un signal au reste du cerveau pour indiquer la direction à suivre. Mais sans dopamine, ce système ne fonctionne pas, et c'est ce qui explique que l'animal ne fasse alors plus l'effort d'aller chercher sa récompense. L'une des principales zones du circuit de récompense, la région sous-corticale appelée nucleus acumbens (A-Coume-Bence), joue un rôle particulièrement central dans ce mécanisme : un rat sans noyau acumbens est incapable de retenir le caractère positif ou négatif d'un indice et n'a donc pas tendance, naturellement, à rechercher les situations associées par le passé à une récompense.
Ces neurones dopaminergiques sont également sensibles à la probabilité de la récompense ou de la punition, et à sa proximité temporelle. Ils s'activent d'autant plus fortement que la récompense prédite est importante, probable ou proche. Si un indice n'est pas systématiquement suivi par une récompense, ou si celle-ci est trop décalée dans le temps, la réaction des neurones à l'indice est faible.
Les neurones dopaminergiques manifestent également leur mécontentement si la récompense obtenue n'est pas celle escomptée, par exemple si le gros paquet sous le sapin n'est pas le vélo tant souhaité : leur activité diminue brutalement si la récompense est plus faible que celle espérée ; en un mot, ils boudent. Ils vont même jusqu'à se taire totalement quand le cerveau est confronté à une stimulation aversive, désagréable. Mais, bons joueurs, ils réagissent aussi vivement si la récompense est plus forte que prévue, ou si un événement inattendu et agréable se produit. Le circuit de récompense est donc capable de prendre en compte dans ses calculs non seulement l'intensité du plaisir que devrait procurer la récompense si elle est obtenue, mais également la proximité dans le temps de cette récompense et les chances de l'obtenir. La grande force des neurones dopaminergique réside donc dans leur capacité d'anticipation, qui leur permet d'orienter le comportement vers les actions et les situations qui récompensent le cerveau... et d'éviter les punitions. Avec ce système, le cerveau dispose donc d'un outil formidable pour évaluer tout ce qui l'entoure en fonction de son utilité potentielle. Nous disposons là d'un petit mécanisme qui note chaque situation en fonction du type de récompense, ou de punition, qu'elle est susceptible d'amener. C'est le système de « Post-it » de l'amygdale, étendu à tout le circuit de récompense.
L'expérience des rats d'Olds et Milner, largement confirmée depuis, nous montre que le cerveau privilégie spontanément les actions stimulant l'activité des neurones du circuit de récompense. Ces actions privilégiées sont celles qui mettent l'organisme dans des situations où il est récompensé, ou simplement au contact d'indices annonciateurs de récompenses. Ce mécanisme impose donc une contrainte très forte sur le comportement : un rat qui a pris l'habitude de recevoir une drogue récréative dans une pièce de son habitat va y retourner, encore et encore, même s'il n'y reçoit plus rien ; car, pour ce rat, le simple fait d'être dans cette pièce suffit à activer le circuit de récompense. Et ce qui est vrai chez le rat l'est aussi chez l'homme : le cerveau d'un cocaïnomane libère de la dopamine quand on lui montre des vidéos du quartier où il se procure sa drogue. Le cerveau humain est donc lui aussi sensible à tous les indices qu'il a pu associer à des sensations de plaisir. La cocaïne ne fait que détourner un système par ailleurs normal et bénéfique ; n'importe quel stimulus ou situation peut causer la libération de dopamine, du moment qu'il a été associé avec une récompense : des lieux, des personnes, voire des sensations corporelles. En s'activant, les neurones dopaminergiques du système de récompense encouragent l'individu à engager, ou poursuivre, tout effort susceptible d'amener une récompense. Même si la libération de dopamine n'est pas le corrélat de la sensation de plaisir, toute action, ou toute perception, capable de la déclencher finit par être recherchée.
Il est alors facile de comprendre comment le circuit de récompense intervient dans la captivation de l'attention. L'orientation de l'attention vers un stimulus apprécié de ce circuit déclenche naturellement la réaction des neurones dopaminergiques ; et la réorientation de l'attention vers un autre stimulus, plus neutre, interrompt cette réponse dopaminergique, comme si ce dernier était associé à une punition. L'attention a donc naturellement tendance à rester fixée sur l'image ou le son « dopaminé », tant que celui-ci stimule le circuit de récompense.
Quand Jenson Button aperçoit la jolie fille sur l'affiche, ses neurones dopaminergiques envoient une petite bouffée de dopamine dans son circuit de récompense pour l'encourager à regarder. Cette capture de l'attention est jugée favorable, car annonciatrice d'une possible récompense. Mais lorsque le pilote détache son regard, ces mêmes neurones réagissent en stoppant net leur décharge, le cerveau est puni. Et ce petit mécanisme se reproduit des milliers de fois par jour, non seulement au volant, mais partout dans notre vie, dès que nous nous détournons d'une perception agréable ; l'attention a naturellement tendance à s'installer, à séjourner – pour reprendre le terme de Damasio, to dwell – sur tout ce qui stimule le circuit de récompense : des objets ou des personnes rencontrés, des situations vécues... mais également des raisonnements ou des rêveries, car le circuit de récompense, surtout celui de l'homme, sait parfaitement réagir à des situations abstraites, imaginées. C'est l'une des raisons pour lesquelles nous aimons tant basculer vers notre monde intérieur, peuplé d'images mentales, de pensées et de raisonnements. Le cerveau se coupe du monde qui l'entoure pour partir dans son petit univers à lui.
ATTENTION
Par Jean-Philippe Lachaux : ("Le cerveau attentif")
Lorsque nous sommes tristes ou angoissés, ces sentiments se nourrissent de notre attention. Sans elle, ils ne sont rien et retournent au néant. L'attention nous permet de souffrir. Mais elle nous permet aussi de comprendre, de nous émerveiller et de savourer le spectacle du monde et de notre existence.
Habitudes attentionnelles
Attention conjointe ou sociale
Face à des stimuli ou à des environnements visuels connus, le cerveau a donc pris l'habitude, parfois depuis l'enfance, de faire attention aux éléments les plus riches en informations. Avec l'habitude, ces déplacements « intelligents » de l'attention finissent rapidement par s'enchaîner de façon automatique, sans effort volontaire, pour former des habitudes attentionnelles.
Capture de l'attention (ou captivation)
Quand cet événement est jugé suffisamment intéressant ou important, la cascade se prolonge pendant un temps plus ou moins long qui est fonction du degré de concentration de la personne à ce moment-là. Pendant cette deuxième phase, l'attention n'est plus disponible pour la tâche principale, qu'il s'agisse de la préparation d'un service au tennis ou bien de mots croisés. C'est une phase de fascination ou, pour utiliser un néologisme, de captivation, pendant laquelle l'attention reste captive de son ravisseur. La force de cette captivation se mesure alors par le temps que met le cerveau pour rediriger l'attention vers son objectif premier, au-delà de la fraction de seconde de la capture initiale.
Certains phénomènes moteurs, émotionnels et cognitifs suivent généralement la capture de l'attention et ralentissent son retour vers son objet principal.
a) - Phénomènes moteurs
La voie "où" et la voie "quoi"
Au sein de la voie dorsale qui traverse le lobe pariétal, les informations spatiales sont traitées par des groupes de neurones surveillant chacun une région précise et fixe de l'espace. Chaque neurone signale par son niveau d'activité la présence, ou l'absence, d'un objet, statique ou en mouvement, dans la portion d'espace visuel qu'il a charge de surveiller. Il suffit donc de mesurer l'activité de ces neurones pour connaître la position, et le mouvement, de tout ce qui nous entoure ; exactement comme on lirait une carte.
L'attention guide l'action
Il s'agit toujours d'une association visuo-motrice, mais qui n'associe plus une position dans l'espace avec un mouvement destiné à l'atteindre, mais un objet, une forme, avec un geste de saisie et d'utilisation. Toutefois, c'est encore le lobe pariétal qui s'en charge, grâce à d'autres neurones situés dans sa partie basse, près du pouce du gant de boxe, dans la partie dite antérieure du sillon intrapariétal : AIP.
Cette région pariétale semble se « souvenir » de la façon dont on utilise un club de golf ou un tournevis, et de ce que l'on fait avec des cacahuètes. Les neurones d'AIP qui réagissent fortement à la vue d'un tournevis réagissent également à la vue d'une clé, mais pas d'un stylo ; parce qu'une clé s'utilise comme un tournevis, en la tournant. La nature faisant bien les choses, AIP se situe à côté des régions du lobe temporal chargé d'identifier les objets dans la voie visuelle « quoi ? ». Après tout, savoir ce qu'est un tournevis, c'est aussi savoir ce que l'on peut en faire. Justement, les patients chez qui AIP est endommagée, souffrent souvent d'un trouble appelé Apraxie idéatoire.
Le lobe pariétal est donc capable d'associer à chaque objet familier le ou les programmes moteurs le plus souvent associés par le passé à cet objet ou à d'autres lui ressemblant. C'est bien le lobe temporal qui reconnaît l'objet, à partir de sa forme, mais c'est le lobe pariétal qui propose les actions à réaliser pour l'atteindre et l'utiliser ; le cortex frontal se charge ensuite de les exécuter ou non. Dans le cas des cacahuètes, le scénario cortical est donc sans doute le suivant : le bol est d'abord identifié et reconnu par les régions visuelles du lobe temporal qui activent immédiatement dans le lobe pariétal les neurones chargés de préparer le geste à réaliser pour attraper les cacahuètes entre le pouce et l'index. Cette activité se propage ensuite vers les neurones du cortex prémoteur, puis moteur, qui déclenchent l'exécution du geste. Pendant ce temps, d'autres régions du lobe pariétal ont déterminé la position du bol dans l'espace, et calculé le mouvement du bras à réaliser pour atteindre le bol avec la main. La convergence de ces deux vagues d'information au niveau du cortex moteur aboutit à l'exécution d'un geste parfaitement coordonné, grâce à l'aide supplémentaire des régions sous-corticales et du cervelet. En suivant ce raisonnement, il apparaît que l'un des rôles majeurs de la partie arrière de notre cerveau, et notamment du lobe pariétal, consiste à proposer des actions aux régions motrices du lobe frontal, en fonction de ce que nous avons sous les yeux, et de nos habitudes. Vous voyez une tasse de café ? Votre « cerveau arrière » vous propose de la boire. Vous voyez une porte ouverte ? Entrez, je vous en prie ! Cette partie arrière du cerveau est donc force de proposition, avec quelques erreurs parfois : vous est-il déjà arrivé, à vous aussi, de sortir vos clés en approchant du portillon du métro ?
L'attention joue à ce stade un rôle essentiel, car ce bel enchaînement entre perception et action se heurte mécaniquement à un goulet d'étranglement : la Nature ne nous ayant dotés que de deux bras et deux jambes, nous ne pouvons pas agir en même temps sur tous les objets qui nous entourent : saisir les cacahuètes, tourner la page du journal, nous gratter le menton et demander l'addition. Nous ne sommes pas des pieuvres. Un mécanisme de sélection est donc fatalement nécessaire pour limiter le nombre de propositions d'action qui atteignent le cortex moteur. Et revoilà l'attention. En sélectionnant un par un les objets qui sont devant nous, l'attention permet d'organiser dans le temps notre interaction avec le monde, une action après l'autre. En ne privilégiant qu'un objet à la fois, elle évite aux propositions d'action de s'accumuler à l'entrée du cortex moteur en le paralysant. En revanche, l'arrivée de l'attention sur un objet déclenche naturellement un ensemble de propositions d'actions relatives à celui-ci, qui expliquent pour une part le processus de captivation motrice qui prolonge la capture.
Fort heureusement, le cortex moteur n'exécute pas toutes les actions que lui propose le lobe pariétal. Il vous arrive rarement, j'imagine, d'enfiler votre manteau sur votre pyjama pour sortir de chez vous, juste parce que vous avez aperçu le portemanteau sur le chemin des toilettes. Mais ce n'est pas, cette fois, à cause du goulet d'étranglement moteur, mais grâce à un garde-fou situé dans le cortex frontal, qui filtre les propositions d'actions jugées trop farfelues. Quand ce garde-fou fonctionne bien, la plupart des propositions du lobe pariétal passent inaperçues et nous n'en avons même pas conscience. Mais il suffit qu'une des composantes du garde-fou déraille pour que la partie arrière du cerveau prenne le pouvoir sur le corps". C'est ce qui arrive aux malheureux atteints de lésions du complexe moteur supplémentaire, ou CMS, un ensemble de régions corticales situées juste en avant du cortex moteur dans le lobe frontal".
b) - Phénomènes émotionnels
L'attention peut donc rester piégée, captive, si la sensation éprouvée au contact du distracteur le justifie. La victime tarde à quitter son ravisseur ; c'est la version neuronale du fameux syndrome de Stockholm, qui décrit le sentiment d'attachement que ressentent parfois les otages pour leurs geôliers. Une nouvelle fois, la capture de l'attention est suivie par une phase de captivation, mais qui n'est pas motrice cette fois, mais émotionnelle, et que Damasio désigne sous le terme d'attention dwelling, qui traduit bien cette idée de durée – to dwell signifie « rester, habiter, résider ». Cette forme de captivation dépend principalement de l'intensité et de la qualité du ressenti, plaisir ou déplaisir, en présence de l'objet, comme si la captivation avait pour but de prolonger cette sensation ou de la réduire.
En français, le mot « distraction » fait d'ailleurs souvent référence à cette notion de plaisir. Se distraire, c'est s'amuser, s'extraire de son travail par exemple, pour passer à une autre activité agréable qui change les idées : « Arrête un peu de travailler, viens te distraire ! » Et il est parfois bien difficile de rester concentré en présence de charmants distracteurs : le champion de formule 1 Jenson Buthon peut en témoigner, qui déclarait après un grand prix avoir été gêné à la sortie d'un virage par une affiche publicitaire montrant un mannequin en tenue légère. Manifestement, le circuit qui permet au cerveau de ressentir du plaisir joue un rôle central dans le phénomène de déconcentration.
La recherche du plaisir est l'un des grands moteurs de la distraction. Par chance, ce circuit du plaisir commence à être bien connu ;
L'expérience d'Olds et Milner révèle l'un des moteurs fondamentaux de la motivation : entre deux actions possibles, l'animal, ou l'homme, choisit spontanément celle qui le récompense le plus, c'est-à-dire celle qui provoque l'activation la plus forte du circuit
de récompense. Pour ces rats, aucune action ne pouvait activer plus
efficacement leur circuit de récompense qu'une stimulation électrique directe, pas même l'ingestion de nourriture. Les rats finissaient donc par mourir de plaisir, malheureusement, mais tout à fait logiquement, comme d'authentiques junkies devenus totalement esclaves de leur drogue. Et ce n'est pas qu'une image : nous savons maintenant que le circuit de récompense joue un rôle central dans le phénomène d'addiction. Pourtant, la vocation première du circuit de récompense n'est pas de favoriser la toxicomanie, mais d'encourager les comportements les plus utiles pour l'animal, ceux permettant de maintenir l'organisme dans sa zone de confort, dans ce que l'on appelle l'état d'homéostasie : manger quand on a faim est agréable tout comme se mettre au chaud quand il fait froid'. Pour l'animal, cette recherche implique de se mettre en quête de nourriture quand son organisme manque de glucose et de se réchauffer quand la température de son corps baisse, etc. : ce qui est utile pour l'organisme est ressenti comme agréable selon le principe d' alliesthésie'. Dans le cerveau plus complexe de l'homme, le circuit de la récompense réagit à des sources de plaisir très variées, comme la consommation de nourriture ou de cocaïne, certes, mais aussi à une bonne soirée entre amis, une partie de jeu vidéo ou un petit moment de détente musicale ; bref, à tout ce qui a une utilité subjective pour l'individu, pour reprendre le terme des spécialistes de la prise de décision.
Le circuit de récompense réagit non seulement aux récompenses, mais à tous les stimuli ayant une valeur hédonique forte, qu'elle soit positive ou négative. Cela ne veut pas dire que le cerveau réagit de la même manière aux récompenses et aux punitions, mais simplement que les neurones qui réagissent aux uns et aux autres sont situés dans les mêmes structures. Dans l'amygdale, par exemple, les récompenses et les punitions activent des populations de neurones différentes, mais adjacentes. C'est aussi vrai dans les autres structures clés du circuit de récompense : le pallidum ventral, le nucleus acumbens, le striatum ventral et le cortex orbito-frontal. La proximité de ces deux populations leur permet de rivaliser entre elles grâce à des mécanismes d'inhibition mutuelle : les neurones « récompense » tentent d'empêcher les neurones « punition » de s'activer, et réciproquement. Le risque d'ambiguïté est donc faible : un stimulus peut difficilement être perçu à la fois comme une récompense et comme une punition. Le cerveau est finalement assez manichéen.
c) - Phénomènes cognitifs
Quand il s'agit d'étudier les pensées spontanées, le phénomène ne peut pas être induit par l'expérience, par définition — on conçoit mal une expérience qui demanderait aux participants de penser spontanément à certains moments et pas à d'autres — et les pensées spontanées ne se traduisent pas non plus par un comportement observable, mesurable, de l'extérieur. À moins d'interroger directement la personne pendant l'expérience — ce qui ne constitue par une mesure objective —, l'expérimentateur n'a aucun moyen de savoir exactement à quels moments les pensées se produisent.
Malgré cette mise en garde, certains chercheurs ont tout de même décidé de franchir le Rubicon pour étudier l'activité cérébrale qui accompagne les pensées spontanées... en demandant de temps à autre au participant s'il est en train de penser. Le débat est alors ouvert : non, il ne s'agit pas d'une mesure objective ; oui, l'expérience fait directement appel à l'introspection des sujets et fait confiance à leurs rapports verbaux ; non, ce n'est pas parfaitement rigoureux ; oui, ces expériences font polémique. Et oui, ces études reposent entièrement sur la capacité du participant à décrire de façon fiable l'état mental dans lequel il se trouve. Malgré tout, des équipes comme celles de Neil MacCrae à Aberdeen, Sam Gilbert à Londres ou Jonathan Schooler à Santa Barbara ont réussi à publier leurs travaux dans les meilleurs journaux, ce qui atteste d'une évolution des mentalités en neurosciences cognitives et d'une prise de distance vis-à-vis du behaviorisme pur et dur.
- SIC et SOC
Réseau par défaut ou énergie noire du cerveau
Le réseau par défaut est donc un ensemble de régions cérébrales actives dès que l'activité cognitive n'est pas contrainte de l'extérieur par une tâche demandant une attention soutenue à l'environnement. Les régions corticales qui le composent sont distribuées à la fois sur la face latérale du cortex, celle que nous voyons en regardant le cerveau de côté, et le long du mur médian séparant les deux hémisphères. Sur la face latérale, le réseau occupe la fameuse jonction temporo-pariétale — souvenez-vous des mots croisés — et la partie basse du cortex préfrontal tout à l'avant du cerveau, dans ce que l'on appelle le cortex préfrontal ventro-latéral, juste devant l'aire de Broca. Le long du mur médian, le réseau s'étend autour du corps calleux, à l'arrière du cerveau, au niveau du cortex cingulaire postérieur, et à l'avant, au niveau du cortex préfrontal médian. Et il faut ajouter à cela la partie médiane du lobe temporal. Ouf ! Ces régions reviendront dans le cours des explications d'ici la fin du chapitre.
Tout porte à croire aujourd'hui que, lorsqu'un sujet cherche à se concentrer sur une tâche, il existe une compétition entre le réseau par défaut et les régions du cerveau chargées de traiter attentivement les informations importantes pour la tâche. Souvenez-vous de l'expérience de lecture dans laquelle les participants devaient lire l'histoire verte et ignorer l'histoire rouge. Dans cette expérience, la lecture attentive des mots verts s'accompagnait d'une interruption très brève de l'activité dans le réseau par défaut, pendant environ deux dixièmes de seconde, au moment où les régions du cortex préfrontal impliquées dans la lecture se décidaient à rentrer en action. Les mots rouges, qui ne déclenchaient pas cette réaction du réseau frontal de la lecture, ne causaient pas non plus l'interruption du réseau par défaut. Dans une autre expérience menée avec Tomas Ossandon, Karim Jerbi et Juan Vidal, nous avons pu montrer que la recherche d'un objet parmi d'autres s'accompagne d'une chute d'activité dans le réseau par défaut, d'autant plus prolongée que la recherche est longue. Ces résultats montrent à quel point la capacité du cerveau à prendre en compte une stimulation extérieure dépend de la faculté de cette stimulation à interrompre l'activité du réseau par défaut. Mais cette interruption n'est jamais très longue : dès l'information traitée, l'activité du réseau par défaut revient en une fraction de seconde à son niveau de base. Ce réseau n'est donc pas très conciliant. S'il daigne s'effacer pour faire entrer un intrus, il revient aussitôt. Le cerveau n'est pas un hall de gare ouvert aux quatre vents.
Le cerveau semble donc pouvoir évoluer selon deux modes : un mode dédié principalement à l'analyse active de l'environnement sensoriel et à l'adaptation du comportement à cet environnement, et un mode dédié à des processus mentaux d'imagerie principalement visuelle, auditive et motrice, pour manipuler des modèles mentaux de la réalité. Dans ce mode interne, nous évoluons dans une version imaginaire du monde. Nous voyons, nous entendons, nous agissons, nous parlons dans ce monde virtuel comme s'il était réel. Sur le plan anatomique, nos relations avec les mondes externes et internes diffèrent sur certains points ; le réseau par défaut est silencieux dans le premier cas et actif dans le second. Mais cette dissociation est loin d'être totale : perception et imagination utilisent en partie les mêmes régions du cerveau, voire les mêmes neurones, comme ce « neurone Simpsons » du LTM.
Il existe donc une compétition dans le cerveau entre le mode réel et le mode virtuel. Et la prise de contrôle du LTM constitue sans doute un enjeu stratégique particulièrement important. Le LTM intervient à la fois pour rappeler des souvenirs de situations passées ou pour imaginer des situations futures ou imaginaires, mais aussi pour former de nouveaux souvenirs. Si je mesure l'activité de votre hippocampe pendant que je vous montre une liste de mots, je peux prédire ceux dont vous allez vous souvenir, seulement grâce à mes mesures. Et si vous n'aviez pas d'hippocampe, vous ne pourriez vous souvenir d'aucun d'entre eux. Alors, que se passe-t-il lorsque votre hippocampe est momentanément indisponible parce qu'il est occupé par votre univers intérieur ? Dans le LTM, mais également dès les cortex sensoriels, l'attention doit opérer une forme d'arbitrage pour privilégier soit l'analyse des stimulations sensorielles provenant du monde extérieur, soit les représentations imagées qui tapissent notre univers intérieur.
On peut légitimement s'interroger sur l'intérêt pour le cerveau de se couper régulièrement du monde pour explorer ses mondes virtuels. Quel est l'intérêt, du point de vue évolutif, dans un monde potentiellement dangereux ? L'imagination est bien sûr utile, pour anticiper des situations et planifier des solutions, mais pourquoi glissons-nous dans nos rêveries au milieu d'une conversation, d'un cours ou d'un match de tennis ? Pourquoi est-il si dur de rester présent ? Une fois encore, personne ne le sait vraiment ; mais nous pouvons tout de même remarquer certains faits. D'abord, le monde n'est pas toujours en perpétuel changement. Face à mon mur, sur mon coussin, le petit univers qui m'entoure est stable. Je peux donc m'en abstraire quelques dizaines de minutes sans courir pour autant de risque insensé. Et même une fois levé, de retour dans la « vraie » vie, et à moins d'être au volant, le monde ne change pas si vite que cela ; soit parce que effectivement il ne se passe pas grand-chose, soit parce que tout se passe « comme prévu » — le cours des choses ne me surprend pas et les seuls événements susceptibles de se produire sont des situations auxquelles je sais faire face, facilement. Assis sur ma chaise lors d'une réunion, les choses suivent leur petit train-train, et même si quelqu'un s'adresse à moi, je peux compter sur un dispositif d'analyse sensorielle pré-attentif efficace et sur mon circuit de veille attentionnel pour rediriger rapidement mon attention vers la réunion, notamment lorsque j'entends prononcer mon nom. Je peux donc basculer dans mes rêveries sans risque ; au pire, je demanderai à ce que l'on répète la question ; et en tout cas, j'en sortirai vivant.
Il n'y a donc pas tant de risque que cela à basculer dans le mode virtuel. Toutefois, cette absence de risque n'explique pas complètement que nous le faisons si volontiers. Est-ce pour reposer le cerveau ? Peut-être, mais le spécialiste du réseau par défaut, Marcus Raichle, en doute ; il rappelle que la consommation d'énergie du cerveau ne baisse que de 1 % lorsqu'il bascule dans cet état de repos. J'avancerai comme autre explication que la vitesse à laquelle notre monde intérieur varie est bien adaptée à la dynamique naturelle de l'attention. Livrée à elle-même, l'attention ne tient pas en place. La dynamique spontanée du regard lors de l'exploration de scènes visuelles, @? raison de trois ou quatre saccades par seconde, est un exemple parmi d'autres qui illustre le caractère naturellement remuant de l'attention. Le flot régulier de la respiration est l'archétype de ce que l'attention déteste : un stimulus stationnaire, aux variations lentes, dont le cerveau sait tout dès les premières secondes. L'enchaînement des pensées, au contraire, n'impose aucune contrainte de rythme à l'attention. Les rythmes des pensées et de l'attention vont même jusqu'à se codéfimir : chaque nouvelle pensée capture naturellement l'attention et se prolonge jusqu'à ce que l'attention la quitte. L'attention est donc peut-être plus facilement captée par des phénomènes mentaux qui se renouvellent à la vitesse qui lui convient le mieux, plutôt que par des événements extérieurs plus statiques.
Je n'irai pas plus loin dans ces spéculations, concernant les mécanismes à l'oeuvre dans le cerveau lorsqu'on le dit « au repos ». La recherche dans ce domaine est difficile en raison du caractère spontané et éminemment subjectif de l'activité mentale qui semble s'y dérouler. Malgré tout, les recherches avancent et, petit à petit, les avancées des neurosciences cognitives laissent entrevoir certaines des briques avec lesquelles le cerveau se crée ce monde imaginaire où il aime tant s'occuper. Mais n'allez surtout pas croire, après ce chapitre, que l'imagerie mentale et la petite voix se résument à l'activité du LTM et du cortex préfrontal. Ces régions ne sont que des noeuds de vastes réseaux interagissant les uns avec les autres. Parce qu'elle implique la manipulation de concepts et de situations, la production du discours interne fait appel à des régions du lobe temporal antérieur et médian impliquées dans la mémoire sémantique et épisodique. Comment parler des Beatles sans activer ces neurones du LTM qui réagissent au simple mot « Beatles » et au visage de Paul McCartney ? Chaque élément du discours mental induit une modification de la représentation mentale que nous avons en tête. En pensant la phrase : « Je dois sortir le chien », vous évoquez naturellement un ensemble de représentations mentales associées à votre chien et au fait de le sortir, comme le trajet que vous empruntez d'habitude. L'évocation de ces lieux au sein de votre univers mental génère un autre discours, etc. Tous ces systèmes agissent donc ensemble pour produire la réaction en chaîne que nous appelons « pensée », et qui correspond à une forme de captivation cognitive de l'attention.
Système exécutif
Il intervient en particulier pour désengager le pilote si celui-ci propose une action inadéquate. La tâche de Stroop, que beaucoup utilisent pour définir l'attention exécutive, est un exemple de situation mettant en jeu le système exécutif, puisque le sujet doit inhiber un comportement habituel mais inadapté : la lecture du mot qui s'affiche à l'écran. Dans l'immense majorité des expériences menées au laboratoire, le sujet reçoit une consigne très précise — « si vous voyez apparaître une image à gauche de l'écran, regardez vers la droite » —, si bien que le système exécutif intervient surtout pour mémoriser les règles de l'exercice et les faire appliquer par les autres régions du cerveau. Au sein du système exécutif, ces consignes sont traduites sous forme de mécanismes neuronaux — « si le système visuel détecte l'apparition d'un stimulus, il faut inhiber l'activité du champ oculomoteur pariétal, et programmer un mouvement d'orientation des yeux vers la position opposée à celle du stimulus dans le champ visuel».
Cette version neuronale des instructions de la tâche porte le nom anglais de task set, que l'on pourrait traduire en français par l'« ensemble de la tâche », qui est un peu ambigu. Le task set est donc une mémoire de toutes les associations stimulus-réponse à mettre en jeu pour réaliser correctement la tâche : si A se produit, alors il faut faire B, et si C se produit, alors il faut faire D. La plupart des activités de la vie courante ont leur task set. Au volant, les stimuli à prendre en compte, c'est-à-dire les informations sensorielles qui décident du comportement, sont surtout la distance aux autres véhicules, les panneaux de signalisation et la courbe de la route ; ainsi, si la route tourne à droite (stimulus), il faut tourner le volant dans le même sens (réponse) et si le véhicule devant nous freine (stimulus), il faut appuyer sur la pédale de frein et rétrograder (réponse).
Le CPFL (cortex préfrontal latéral) est directement connecté aux cortex prémoteur et moteur, si bien que ses neurones sont parfaitement capables de transmettre un signal aux régions motrices pour déclencher la réponse du singe lorsque les figures apparaissent à l'écran, en fonction de La règle que doit suivre l'animal. Lorsque c'est un triangle qui apparaît à l'écran, ces neurones n'envoient aucun signal vers le cortex moteur. Et si la règle change et que le singe doit maintenant appuyer sur Le levier quand un triangle apparaît, ces neurones du CPFL cessent de s'activer, et d'autres neurones voisins prennent le relais, des neurones « si triangle, alors appuyer sur levier ». Ces autres neurones réagissent exactement comme Les premiers, mais avec cette fois une préférence pour les triangles. Sur le plan comportemental, le singe ne réagit plus aux ronds, car les neurones « si rond, appuyer sur levier» ne sont plus actifs. Grâce à ce mécanisme simple, le CPFL peut facilement s'adapter à un changement de consigne en inhibant les neurones « si rond, alors... » et en activant les neurones « si triangle, alors... ». L'animal se souviendra de la consigne et continuera à l'appliquer, tant que l'activité des neurones correspondant à cette consigne se maintiendra dans le CPFL. C'est simple, mais il fallait y penser. En observant ces neurones, il est donc possible de connaître la règle que Le singe est en train d'appliquer.
La programmation et le maintien en mémoire d'un task set demandent un effort, car il faut maintenir actif, de façon continue, un ensemble de neurones du cortex préfrontal. Chez un animal peu motivé, l'activité retombe rapidement et le cerveau revient rapidement à ses vieux automatismes. Masataka Watanabe et son équipe à Tokyo s'en sont aperçus en enregistrant l'activité du CPFL chez des singes tout en faisant varier la récompense reçue après chaque essai réussi', dans une tâche semblable à celle des ronds et des triangles. L'activité des neurones du CPFL chargés de garder en mémoire la consigne était beaucoup plus forte quand les singes recevaient du raisin, leur récompense favorite, que des pommes de terre, un mets qu'ils n'aiment pas trop. Un singe plus motivé est donc plus difficile à distraire de sa tâche, car l'activité neuronale dans le CPFL est plus forte.
Sortir des sentiers battus : le système exécutif face aux habitudes
DÉSIR MIMÉTIQUE
Le philosophe René Girard, né en 1922, est l'inventeur de la fameuse théorie du désir mimétique, qui a jeté les bases d'une nouvelle anthropologie, selon laquelle la pulsion motrice et créatrice de l'humanité est fondée sur une spirale compétitive : le désir du même. Cette spirale serait mortelle pour le genre humain, si elle n'était canalisée par les rituels religieux. Cette théorie, d'abord littéraire (fondée notamment sur l'étude des écrivains romanesques : Cervantès, Stendhal, Dostoïevski, Proust), puis philosophique, a trouvé un renfort inespéré dans la neuropsychologie et la découverte des neurones miroirs. De quoi s'agit-il ?
Deux bambins dans un bac à sable se disputent férocement un seau en plastique rouge. Vous tentez de les raisonner, leur montrant d'autres jouets, bien plus beaux. Mais rien à faire, ils veulent le même ! Il faut les séparer, écumant, hurlant. S'ils en avaient le pouvoir, sûr que chacun anéantirait l'autre dans un éclair de violence. Prenez maintenant un couple. Depuis quelque temps, cet homme ne regarde plus sa femme qu'avec ennui, il ne la désire plus. Survient un étranger, dont les yeux brillent quand il devient ravissante. En peu de temps, la flamme du mari renaît. Hier indifférent, il serait soudain prêt à se battre pour réaffirmer son « amour éternel» à son épouse. Ainsi ne désirons-nous rien tant que ce que désire l'autre. Éros, pulsion de vie et de création, fonctionne en miroir nous désirons ce que nous désigne le désir d'autrui.
Étendu à l'humanité, le désir mimétique, analyse René Girard, entraînerait tout dans sa violence, si celle-ci n'était focalisée, à intervalles réguliers, sur un bouc émissaire, une victime expiatoire et sacrée, qui prend sur elle la rage collective de ce désir, avant de disparaître. D'où, sans doute, le fait que « sacré » et « sacrifice » ont la même racine et constituent l'origine de toute culture.
Vidéo René Girard : Le désir est un triangle car le désir subjectif ou objectif est toujours une ligne droite ; on part de l'objet pour aller au sujet ou on part du sujet pour aller à l'objet. On désire un objet parce que celui-ci est meilleur que les autres ou c'est le sujet qui dit : « moi, je désire ce que je veux, et j'ai élu, moi, cet objet là » (je pense que cela vient de mon moi profond). Mais, entre le sujet et l'objet, il y a un médiateur qui est capital (mais que l'on refuse de voir) et qui est modèle. S'il est trop proche, il devient obstacle (rival) ; et cette distance est très importante parce que c'est la distance entre Don Quichotte qui rencontre les moulins à vent sur sa route mais pas de médiateur (le chevalier Amadis de Gaule) et il est donc essentiellement heureux mais lorsque l'on affronte directement le médiateur et que c'est un échec on est malheureux (comme Sanson Carrasco qui affronte Don Guichotte).
Le monde moderne est un monde de rivaux malheureux parce que ce sont des rivaux dans le monde des affaires (pas dans la chevalerie ou des buts lointains et métaphysiques) mais dans le concret ou l'on trouve toujours un rival plus heureux. Plus le médiateur se rapproche, plus les fruits du désir triangulaire se font amers, parce que les fruits du désir triangulaire ne sont plus que la rivalité.
Si on possède l'objet, il n'y a plus de désir et donc déception totale ou alors il y a frustration si l'on ne possède pas l'objet (on reste la langue pendante) car le rival cherche autant à nous humilier qu'à posséder l'objet.
Conception du désir-1 -
Conception du désir-2 Pierre Pachet (professeur des universités, écrivain et essayiste français, né en 1937 de parents juifs d'origine russe ) : La grande pensée du désir fut longtemps celle de la psychanalyse (le triangle oedipien) et tout à coup arrive celle de René Girard, tout à fait sidérante, qui nous dit que le désir est orienté, déterminé et même dicté par la figure de l'autre qui est soit un médiateur externe (éloigné : Georges Clooney et le café) soit un médiateur interne (extrêmement proche : parent, ami, rival...)
Dans le désir, peut importe l'objet, peut importe la personne ; ce que nous désirons par dessus tout, ce qui donne naissance et sens au désir lui-même c'est la trajectoire que suit le désir de l'autre que nous envions. Si le désir a une forme ce n'est donc pas celle du coeur mais celle d'un triangle : on ne désire jamais que ce l'autre désire..... Les publicitaires ont compris que, plus qu'un objet, ce que nous désirons par-dessus tout, c'est de faire comme les autres ; mais pas n'importe quels autres ! Ceux que nous admirons, que nous envions, dont nous sommes jaloux, ceux dont le simple regard tourné vers un objet suffit à nous rendre cet objet désirable. C'est cette dimension mimétique du désir qui fait de nous des êtres régis par le désir de copier, d'égaler, de dépasser, bref qui fait de nous, tous autant que nous sommes des snobs.
Jean-Michel Oughourlian (neuropsychiatre) : La grande sagesse c'est de reconnaître que notre désir ne nous appartient pas. Il faut reconnaître modestement que l'on n'est pas maître de son désir et que finalement on ne désire que ce que l'autre désire (on n'est que les autres dit Laborit) et çà c'est une profonde sagesse qui amène à un apaisement complet ; mais c'est à la portée de très peu de gens !
René Girard à non seulement bien vu cette imitation mais il a également été un pas plus loin en voyant que si le désir n'est pas partageable, c'est la naissance de la rivalité par l'envie (on devient envieux). Nous passons alors de l'imitation simple qui n'est autre que l'apprentissage (apprendre à parler, à écrire etc..) à quelque chose qui est beaucoup plus fort et qui est l'envie (la jalousie, le ressentiment) ; le désir rival peut alors mener à l'élimination, à la destruction du rival (ou des rivaux).
Le désir peut également provenir pour chacun de nous à travers la culture dans laquelle nous baignons et des modèles qui nous imprègnent et que nous intégrons inconsciemment. Le désir obstacle ou interdit est également très attirant ! Il y a donc le désir horizontal (désir provenant d'un médiateur) et le désir vertical (celui provenant de la culture)
La liberté n'est pas un acquis chez l'homme ; l'homme est en quelque sorte en capacité de se libérer progressivement des mécanismes mimétiques dont il est la victime, des mécanismes rivalitaires dont il est la victime, des mécanismes de jalousie, d'envie, de violence dont il est la victime de manière à progressivement se libérer de tout ce qui finalement fait son malheur...
Les sciences cognitives confirment la dimension mimétique du désir (mai 2012)
Des chercheurs de l’Inserm ont souhaité décrypter les mécanismes cérébraux à l’origine de ce phénomène. Pour cela, ils ont étudié les comportements et l’activité cérébrale de 116 adultes âgés de 18 à 39 ans. Ils les ont installés dans des appareils d’IRM et ont diffusé des dizaines de petites vidéos présentant des objets différents, vêtements, accessoires, aliments, etc, convoités ou non. Par exemple, une part de gâteau montrée du doigt par un acteur alléché et la même part de gâteau délaissée sur une table dans une autre vidéo. A chaque fois, le sujet devait indiquer une "note de désirabilité" allant de 1 à 10 pour l’objet en question. Les résultats montrent que les objets convoités par d’autres, obtiennent de meilleures notes que ceux auxquels personne ne s’intéresse.
Grâce à l’imagerie par résonance magnétique (IRM), les chercheurs ont observé en parallèle l’activité cérébrale des sujets soumis à ces tests. Ils ont constaté deux phénomènes déjà connus. Le premier est l’activation d’une zone appelée système des neurones miroirs quand un sujet observe une action. « Cette région s’active dès qu’un individu fait un geste ou voit quelqu’un d’autre l’effectuer. Cela permet de comprendre l’action d’autrui », explique Mathias Pessiglione (Inserm U975), coauteur des travaux. Le second est l’activation d’une autre zone appelée système cérébral des valeurs si ce même sujet observe un objet séduisant.
Mais les auteurs ont constaté un troisième phénomène. Celui-là nouveau. Quand un participant observe un acteur voulant se procurer quelque chose, l’allumage du système des neurones miroirs stimule le système cérébral des valeurs. Par conséquent, la valeur accordée à l’objet augmente et le désir de l’obtenir également. « Le fait d’observer un individu voulant se procurer un objet augmente son propre désir pour cet objet », résume Mathias Pessiglione.
Ces travaux montrent bien que le désir est contagieux et à l’origine d’un mécanisme cérébral. « A ce titre, un dysfonctionnement des zones impliquées pourraient expliquer certains problèmes de sociabilité, par exemple chez des enfants autistes qui n’accordent pas la même valeur aux objets que les autres ou ne s’intéressent pas aux mêmes choses. Mais cela reste à démontrer », illustre Mathias Pessiglione.
DOULEUR
LA NEUROMATRICE DE LA DOULEUR : Quand on se cogne un orteil sur une roche, on ressent une douleur à cet endroit précis de notre corps. Cette douleur est souvent si vive et bien localisée qu’on a presque l’impression que c’est l’orteil qui a mal. Bien sûr il n’en est rien. Les fibres nociceptives de l’orteil transmettent d’abord leurs influx nerveux à la moelle épinière qui les transmet à son tour au cerveau. Et c’est l’activité nerveuse de certaines régions du cerveau qui va alors nous faire ressentir de la douleur à cet endroit précis. Puis nous faire crier de gros mots, frotter notre orteil, l’éloigner de la source du danger, etc.
Quelles sont ces régions et comment collaborent-elles pour nous faire ressentir les nombreuses propriétés de la douleur (localisation, intensité, type de douleur, charge affective négative, etc.) ? Voilà des questions complexes qui ont été grandement débattues au cours des dernières décennies et qui le sont d’ailleurs encore aujourd’hui.
Une chose semble toutefois certaine : il n’y a pas de « centre de la douleur » unique dont la seule activité pourrait rendre compte de toutes les facettes de la douleur. En d’autres termes, aucune lobotomie d’une région particulière du cerveau ne fait disparaître complètement la douleur.
Cela dit, les expériences menées grâce à des techniques comme l’imagerie cérébrale montrent clairement que de nombreuses régions précises du cerveau modifient leur activité quand on ressent la douleur. Ces régions sont reliées entre elles et forment un réseau que certains appellent la « matrice de la douleur ». Et selon ce que l’on connaît déjà de ces régions, on les associe souvent à différents aspects de la douleur.
C’est le cas de la formation réticulée du tronc cérébral, l’une des premières structures cérébrales à recevoir des connexions des voies ascendantes de la douleur en provenance de la moelle épinière. L’activation de la formation réticulée contribue aux réactions d’éveil et de vigilance associées à la douleur. Ses neurones peuvent agir sur le rythme cardiaque, la pression artérielle, la respiration et d'autres fonctions vitales que la douleur peut affecter. C’est aussi la formation réticulée qui fait qu’une douleur peut passer inaperçue si notre attention est focalisée sur une tâche captivante.
Les voies ascendantes de la douleur vont ensuite faire escale dans le grand relais sensoriel qu’est le thalamus. Elles y feront des connexions dans différentes sous-régions du thalamus, notamment dans le noyau ventral postérolatéral (ou VPL) situé dans la partie ventrale, postérieure et latérale du thalamus.
Le noyau VPL joue un rôle majeur dans la discrimination sensorielle de la douleur. En effet, ses neurones projettent ensuite leur axone au cortex somatosensoriel reconnu pour sa capacité à localiser la douleur et à en évaluer l’intensité. C’est aussi dans ce noyau VPL que passent les messages du toucher avant de se projeter sur le même cortex somatosensoriel. Les informations relatives au toucher et à la douleur sont tout de même séparées dans des sous-régions distinctes du noyau VPL.
La partie médiane du thalamus reçoit pour sa part des connexions de la formation réticulée et démontre une organisation somatotopique (voir encadré) plus ténue qui l’éloigne de la fonction discriminative. Avec ses neurones qui font des connexions dans le lobe frontal sur les aires corticales motrices, cette région médiane du thalamus participe à l'élaboration des réactions motrices et émotionnelles liées à la douleur.
Les noyaux intralaminaires du thalamus, situés tout près de la région médiane, font aussi partie de cette zone dite « non spécifique » du thalamus qui participe à la réponse d’éveil à la douleur. En plus du lobe frontal, les noyaux intralaminaires vont rejoindre différentes régions du système limbique. Et comme le cortex frontal envoie lui-même de nombreuses projections vers le système limbique, on a ici affaire à coup sûr à un système impliqué dans la composante émotionnelle désagréable de la douleur et à la réponse comportementale destinée à l’amoindrir.
L’abondance de ces connexions réciproques que l’on retrouve dans le cortex cérébral y rend le message nociceptif beaucoup plus difficile à suivre. Les études d’imagerie cérébrale nous permettent néanmoins d’établir une implication des cortex somatosensoriels primaires, secondaires, cingulaires antérieurs et insulaires dans la perception de la douleur.
Après avoir rejoint la formation réticulée du tronc cérébral, puis certains noyaux du thalamus, le message douloureux atteint le cortex cérébral.
Les outils d’imagerie cérébrale permettent donc de faire des associations entre des structures cérébrales particulières et les différentes dimensions de ce phénomène complexe qu’on appelle couramment la douleur. Ensemble, ces régions cérébrales interconnectées entre elles forment ce qu’on appelle la matrice de la douleur. Ses différentes composantes peuvent être associées spécifiquement, mais pas exclusivement, à l’anticipation de la douleur, à sa discrimination ou à ses manifestations affectives désagréables, comme c’est le cas pour le cortex cingulaire antérieur.
On explique cette association entre les affects négatifs de la douleur et l’activité du cortex cingulaire antérieur par le fait que celui-ci intègre les inputs sensoriels au traitement cognitif. Cela permet de produire une réponse motrice appropriée à la stimulation douloureuse, comme des comportements d’évitement. L’émotion étant liée à la motivation et celle-ci à l’action, on comprend l’importance du cortex cingulaire antérieur dans les réactions affectives à la douleur qui nécessitent une réponse comportementale immédiate.
Et si c’est la partie antérieure du cortex cingulaire qui est la plus souvent citée quand on parle de la douleur, les travaux de Burkhart Bromm montrent que c’est d’abord le cortex cingulaire postérieur qui répond le premier au message nociceptif (environ 220 millisecondes après la stimulation nociceptive). Cette activité se déplace par la suite vers les parties médianes et antérieures du cortex cingulaire, avant de s’éteindre dans le cortex frontal autour de 300 millisecondes après le début du stimulus.
C’est aussi le cortex cingulaire postérieur qui permettrait de fondre en une perception unifiée l’affect négatif de la douleur à sa localisation, sa nature et son intensité grâce à ses connexions au cortex pariétal reconnu pour son rôle dans l’intégration de modalités sensorielles.
La partie postérieure du cortex pariétal intervient également dans l’attention au stimulus douloureux, de même que la région dorsolatérale du cortex préfrontal droit qui fait aussi partie de ce réseau cortical attentionnel. On sait en effet à quel point le détournement de l’attention du stimulus douloureux peut diminuer la sensation subjective de douleur, et comment ce bien-être ressenti s’accompagne d’une diminution réelle d’activité dans des régions cérébrales associées à la douleur.
Le cortex préfrontal n’est pas seulement impliqué dans les fonctions dites «supérieures» impliquant souvent l’attention, mais aussi dans l'apprentissage des sensations nociceptives, et donc dans le développement d’un affect négatif associé à ces situations. Il est donc extrêmement bien placé pour avoir son mot à dire sur l’anticipation et le contrôle de la douleur.
Par exemple, lors d’expériences conçues pour inciter le sujet à anticiper la venue de chocs électriques douloureux, ceux-ci étaient appliqués dans deux conditions : sans crème, et avec une crème placebo présentée comme analgésique. Durant la condition placebo, outre le fait que les sujets rapportent une douleur moindre, on observe une diminution concomitante de l’activité nerveuse dans des régions associées à la douleur comme le thalamus, le cortex somatosensoriel primaire et secondaire, le cortex cingulaire antérieur et le cortex insulaire (ou insula). Cependant, l'anticipation d’une douleur moindre dans cette condition placebo amène au contraire une augmentation d’activité électrochimique dans le cortex préfrontal ainsi que dans une région du mésencéphale incluant la substance grise périaqueducale.
Le cortex préfrontal étant également associé à certaines formes de mémoire de travail, autrement dit au maintien temporaire d’idées, d’informations ou de pensées en vue d’un contrôle cognitif, on voit comment tout cela pourrait lui permettre de jouer un rôle dans l’anticipation d’un soulagement à l’origine de l’effet placebo.
Quant à la substance grise périaqueducale, son activation en parallèle avec le cortex préfrontal durant l’anticipation d’un soulagement va dans le sens de l’hypothèse déjà avancée voulant que des mécanismes préfrontaux déclenchent la libération d’opioïdes endogènes dans la substance grise périaqueducale durant l’effet placebo. De plus, ce noyau du mésencéphale reçoit de l’information de nombreuses structures cérébrales liées à l’intégration des processus émotionnels.
La région de la substance grise périaqueducale reçoit aussi des afférences des fibres nociceptives ascendantes susceptibles elles aussi de déclencher les mécanismes de contrôles descendant qu’exerce cette région sur les neurones de la corne dorsale de la moelle épinière.
On sait maintenant que cette analgésie endogène peut être déclenchée par la stimulation de plusieurs autres structures sous-corticales, allant du bulbe au diencéphale. C’est le cas notamment du noyau du raphé (l’un des plus efficaces avec la substance grise périaqueducale), du noyau réticulaire latéral, du noyau du tractus solitaire, du locus coeruleus, de l’aire parabrachiale et de hypothalamus latéral.
D’autres structures sous-corticales contribuent à différents phénomènes associés à la douleur. Ainsi, l’envoi d'informations nociceptives de la formation réticulée et du thalamus non spécifique à la structure régulatrice végétative par excellence qu’est l’hypothalamus sera à l'origine de l’augmentation de la sécrétion des hormones de stress et de l’activation du système nerveux sympathique. Les mêmes projections, en activant le striatum, favoriseront les réponses motrices d’alarme en grande partie automatiques déclenchées par une stimulation douloureuse.
Les interconnexions importantes entre le cortex cingulaire antérieur et l’amygdale, un haut lieu de la régulation viscérale émotionnelle, expliquent la sudation, l’accélération du rythme cardiaque, l’augmentation de la tension artérielle ou les nausées provoquées par une douleur intense.
Finalement, la localisation anatomique particulière de l’insula, ainsi que ses liens privilégiés avec le système limbique en font un candidat idéal pour servir d’interface entre l’information sensorielle en provenance du corps et l’état cognitif particulier d’une personne à un moment donné. Car une sensation subjective comme la douleur est construite justement par l’intégration de cette information sensorielle et cognitive. Or l’insula (plus spécialement l’insula antérieure droite) est l’une des structures cérébrales les plus fréquemment activées non seulement directement par un stimulus douloureux, mais aussi lorsqu’une personne regarde des images de situations douloureuses et imagine que c’est elle qui les subit.
Les recherches portant sur les bases neuronales de l’empathie révèlent ainsi une superposition partielle des aires cérébrales actives lors d’une douleur subie versus une douleur observée chez quelqu’un d’autre, superposition qui comprend l’insula et le cortex cingulaire antérieur. La vue d’une image évoquant la peur, un autre affect négatif proche de la douleur, entraîne pour sa part l’augmentation d’activité dans le cortex cingulaire antérieur et dans des structures comme l’amygdale, mais pas dans l’insula. Si la peur et la douleur provoquent tous les deux un état émotionnel désagréable associé à une réaction de retrait et de protection, on voit que leurs bases neurologiques ne sont encore une fois que partiellement superposables.
On découvre aussi que l’activité dans ce réseau est très sensible aux processus de régulation « de haut en bas » (« top down », en anglais) ce qui expliquerait des phénomènes comme l’effet placebo. Sans compter l’intégration d’une douleur particulière avec notre expérience personnelle et notre héritage culturel qui étend encore davantage le spectre des zones cérébrales impliquées.
Le développement de la théorie du portillon à partir des années 1960 et par la suite de la théorie de la neuromatrice s’appuie sur le constat que la douleur résulte d'une multitude d'interactions et d'échanges d'informations à plusieurs étages du système nerveux. Et la modulation de l’information nociceptive ascendante se fait à chacun de ces multiples relais avant d’être intégrée comme une perception douloureuse. Ce modèle circulaire de la douleur permet surtout de mieux comprendre comment les composantes nociceptives, discriminatives, affectives et comportementales peuvent s'influencer mutuellement.
Le concept de neuromatrice a été mis de l’avant par Ronald Melzack à la fin des années 1980 pour tenter d’expliquer l’étrange phénomène des douleurs aux membres fantômes. Il est en effet très fréquent que des personnes amputées ressentent des douleurs bien réelles qui leur semblent provenir du membre amputé. Ce phénomène illustre clairement que la douleur n’est pas générée par un système à sens unique. Pour l’expliquer, Melzack propose que c’est l’activité nerveuse dans un réseau comprenant plusieurs structures cérébrales qui générait la douleur. Et ce réseau pouvait même générer de la douleur sans qu’il n’y ait de stimulus sensoriel déclencheur.
Dans le cas des douleurs fantômes, un conflit entre la rétroaction visuelle et les représentations proprioceptives du membre amputé pouvait induire dans la neuromatrice une confusion génératrice de douleur. L’utilisation d’un miroir pour donner l’illusion visuelle au patient qu’il a par exemple à nouveau sa main amputée s’est d’ailleurs montrée efficace pour apaiser certaines douleurs fantômes.
Cette matrice de la douleur, ou neuromatrice, constitue donc l’ensemble des régions du cerveau dont l’activité varie lors d’une expérience douloureuse. C’est un vaste espace neuronal où peuvent être codés différents types de douleur. Chacune de ces douleurs singulières auront ce que Melzack appelle une «neurosignature» particulière, c’est-à-dire un pattern d’activation unique de la neuromatrice ou d’un sous-ensemble de celle-ci.
D’autres emploient l’expression d’assemblée de neurones pour décrire ce type d’association neuronale. Et comme les connexions des cerveaux de chaque individu sont différentes dans le détail, les neurosignatures de chaque individu sont forcément différentes. De même, les connexions synaptiques étant modifiables avec l’expérience, une neurosignature va être structurellement différente dans un même cerveau avec le temps qui passe.
Pour tenir compte de toutes les facettes du phénomène des douleurs fantômes, Melzack a proposé une neuromatrice comprenant de nombreuses structures cérébrales impliquées tant dans l’aspect discriminatif, affectif, cognitif et moteur de l’expérience douloureuse des amputés.
Sa neuromatrice incluait au moins trois circuits neuronaux majeurs dont l’importance a été confirmée par les nombreuses études d’imagerie cérébrale qui ont suivi. D’abord une voie nociceptive ascendante spinothalamique latérale discriminative, qui comprend les noyaux ventropostérieurs du thalamus et le cortex somatosensoriel. Ensuite une voie spinothalamique médiane plutôt affective et motivationnelle impliquant le tronc cérébral, les noyaux ventro-médians du thalamus, le système limbique et le cortex frontal). Et finalement des régions associatives du cortex pariétal inférieur.
Le tableau s’est aussi enrichi de régions comme le cortex orbitofrontal, préfrontal (dans les aires de Brodmann 9, 10, 44), moteur (comme l’aire 6 de Brodmann et le cortex moteur supplémentaire), sans oublier certaines régions du mésencéphale comme celle de la substance grise périaqueducale et du noyau lentiforme (ou lenticulaire).
Des régions comme le cortex cingulaire antérieur et l’insula sont même devenues aux yeux de plusieurs des régions clé dont l’activation va nécessairement de pair avec certaines facettes de la douleur, notamment sa composante affective. Sans revenir en arrière et en faire des « centres de la douleur », les neurones de ces régions montrent une grande spécificité à certains aspects de la douleur. Ce qui montre que la neuromatrice de la douleur peut avoir des « nœuds » dont l’activité est plus significative que d’autres.
ÉMOTION / SENTIMENT
Par Antonio Damasio :
Les émotions définies par Damasio incluent donc deux aspects de l’affectivité : l’arousal (excitation, éveil) et l’expression. Aspect expressif, les émotions sont l’action et le mouvement visible pour autrui lorsqu’elles se manifestent sur le visage, dans la voix et à travers les comportements. Aspect arousal, les émotions sont les phénomènes neurophysiologiques, observables empiriquement par les moyens scientifiques. Les émotions ainsi définies sont en principe publiques et observables à la troisième personne, et peuvent être déclenchés et exécutés à l’insu de l’agent de l’émotion. Elles sont avant tout d’ordre non conscient.
Les sentiments ont d’ailleurs les caractéristiques d’être privés, subjectifs et conscients, comme toutes les autres formes de représentations mentales, telles que la pensée, l’information, le savoir ou la connaissance. Les représentations mentales sont privées, parce que seul celui qui le possède peut le voir, comme un film projeté à l’intérieur du cerveau. Les sentiments ainsi définis par Damasio sont les produits issus de l’étape plus élaborée des processus de traitement émotionnel. Ils sont les émotions rendues conscientes. Autrement dit, au stade du sentiment, les émotions, à l’origine somatiques et aveugles, sont éventuellement transformées en images mentales qui nécessitent l’intervention des processus cognitifs de haut niveau.
Interview en 2004 :
Exactement. Même les organismes très simples éprouvent des émotions, c'est-à-dire des réactions naturelles, automatiques, qui les conduisent, directement ou indirectement, à préserver leur corps et à assurer son équilibre interne. Face à une menace, par exemple, un animal va éprouver de la peur et se mettre en retrait. Avant même qu'il fuit ou qu'il se fige, il se produit des changements dans son organisme: la distribution du flux sanguin se modifie, des hormones sont sécrétées... C'est cette série de réactions, visibles ou non, qui constitue ce que l'on appelle «émotion».
L'émotion, au sens le plus simple, serait donc d'abord utilitaire, elle aurait pour but premier de protéger l'organisme, de faire en sorte qu'il se perpétue.
Il n'y a aucun doute qu'elle sert à cela. Spinoza le disait déjà: il y a un désir de durer dans nos corps. On peut dire que ce désir est inscrit dans tous les systèmes biologiques. Tous les animaux ont les mêmes types de réaction face à une menace : la fuite, l'immobilité ou l'agression... Même un organisme unicellulaire comme la paramécie, qui n'a ni corps ni cerveau, s'enfuit lorsqu'il rencontre un danger dans son environnement, une brusque variation de température, une vibration ou le contact d'un objet qui pourrait briser sa membrane.
Mais est-ce vraiment une émotion ?
Il y a là l'essence du processus émotionnel. Les animaux disposent d'une gamme d'émotions primaires: la peur, le bonheur, la tristesse, la colère, la surprise, le dégoût... Ce dernier, par exemple, permet à l'animal de rejeter une protéine qui n'est pas bonne pour lui. Si on mange quelque chose d'avarié, on a immédiatement une réaction de rejet, notre visage se déforme et nous recrachons la substance ou nous la vomissons si nous l'avons déjà ingérée. Le corps se défend. L'animal, dégoûté, agit de la même manière. La mouche, qui possède un système nerveux minuscule, éprouve elle aussi des émotions : si on l'irrite, elle se met à voleter dans tous les sens pour éviter d'être écrasée. Prenez l'aplysia, un petit escargot de mer. Si vous le touchez, il se rétracte, son coeur bat beaucoup plus rapidement, sa pression sanguine augmente, on observe des émissions d'hormones dans tout son corps, et l'animal émet de l'encre noire pour masquer sa présence face au prédateur.
Ne me dites pas qu'il a peur !
Mais si ! On assiste là à un miniconcert de cette émotion qu'on appelle en effet «peur» et qui nous concerne tout autant que cet escargot. Mais il y a une autre question: est-ce que l'aplysia ressent la peur ? Là, j'en doute. Il ne se représente pas cette émotion, il ne la pense pas, à la différence des espèces animales complexes qui ont aussi une gamme d'émotions sociales: la sympathie, l'embarras, la honte, la culpabilité, l'orgueil, l'envie, la gratitude, l'admiration, l'indignation, le mépris... Les oiseaux, les chiens, les singes, les humains ressentent l'émotion, c'est-à-dire qu'ils ont, eux, la possibilité d'établir une relation entre la réaction automatique de leur organisme et l'objet, l'événement, la personne qui en est à l'origine.
C'est ce que vous appelez le «sentiment», qu'il faut donc distinguer de la simple émotion. Peut-on dire que le sentiment est la perception de l'émotion ?
Absolument. Mais c'est aussi la perception de la cause de cette émotion. Lorsque nous éprouvons de la tristesse, nous la percevons physiquement, mais nous avons aussi conscience de ce qui l'a suscitée: une mauvaise nouvelle, la perte d'un objet, la disparition d'un être cher. Les émotions sont des manifestations visibles ou détectables dans le corps (par dosage d'hormones ou par enregistrement des ondes); les sentiments, eux, sont des images mentales, donc cachées... Ce sont en quelque sorte des idées du corps, la conscience d'un certain état du corps lorsque celui-ci est perturbé par un processus émotionnel. Les deux - émotion et sentiment - sont intimement liés, et nous avons tendance à les confondre. Toutes les émotions peuvent devenir des sentiments à partir du moment où nous établissons cette relation de cause à effet entre les transformations de notre corps et ce qui les a suscitées.
Votre escargot marin préféré connaît peut-être la peur, mais il n'a donc pas de sentiments.
Il connaît des émotions, la peur, la sensation de bien-être quand il mange, l'impression que la température ambiante lui convient ou qu'il se sent en sécurité. Il adopte un comportement que l'on peut associer à la souffrance, mais je doute en effet qu'il souffre comme nous ou comme des animaux complexes.
Comment le savez-vous ?
Là, nous sommes dans le domaine de la spéculation, mais je m'appuie sur un constat: notre escargot n'a pas les dispositifs anatomiques nécessaires pour percevoir les changements qui se produisent dans son corps. Pour savoir si tel ou tel animal éprouve ou non des sentiments, il faut aller voir s'il existe, dans son cerveau, une cartographie de son organisme. C'est le cas chez l'homme: les différentes parties de notre corps sont représentées, un peu comme s'il y avait des cartes de correspondance. L'émotion peut aller directement au corps - c'est le cas de l'escargot - susciter des réactions chimiques, des signaux dans les muscles, les viscères. Mais elle peut aussi, c'est la grande différence, partir du cerveau lui-même, agir sur ces cartes de représentation du corps, et simuler en somme un état virtuel.
Quelles sont les espèces dotées d'une aptitude «sentimentale» ?
Je pense que les oiseaux et les mammifères ont des sentiments. Je serais en tout cas navré d'apprendre qu'ils n'en ont pas. Les comportements de ces animaux sont très proches de ceux des humains. Ils disposent des fameuses structures cérébrales qui font l'encartage du corps.
Dans une bonne logique darwinienne, les sentiments auraient donc eux aussi été inventés par l'évolution, au stade des animaux les plus évolués. Mais pour quelle raison? En quoi apportaient-ils quelque chose de plus ?
La nécessité, dans l'évolution, c'était d'avoir une représentation cérébrale du corps. Le cerveau reçoit en permanence des signaux de l'organisme tout entier, qui l'informent sur son état, via les nerfs, le réseau sanguin, et il réagit par des substances chimiques (quand le taux de glucose baisse, on ressent le besoin d'aller manger). Or, dans un cerveau complexe, où les informations reçues sont innombrables, ce système de détection a besoin d'une carte qui représente l'organisme, comme le schéma électrique d'un immeuble. C'est un avantage, car cela permet de répondre immédiatement et précisément aux demandes de l'organisme. L'évolution a donc retenu ce dispositif pour les animaux supérieurs... Mais celui-ci permet aussi d'établir une relation entre une réaction automatique du corps (l'émotion) et ce qui la provoque. La conséquence inattendue, ce sont les sentiments.
Qui seraient donc nés un peu par hasard.
Exactement. Les sentiments ont permis d'aller encore plus loin: un être «sentimental» peut mettre en mémoire différents épisodes émotionnels avec leur cause, il peut prévoir qu'un certain événement risque de provoquer une mauvaise émotion, il échappe à la tyrannie de l'automatisme et acquiert un certain sens du bon et du mauvais. Cela plaide d'ailleurs en faveur de l'existence de sentiments chez les animaux complexes: ils montrent à l'évidence une certaine aptitude morale.
Morale ! Les animaux ?
Si un chimpanzé voit l'un de ses congénères exprimer une douleur, il va éprouver de la compassion à son égard. Il le fait même à l'égard des enfants humains. Récemment, dans un zoo de Chicago, un petit enfant est tombé dans la fosse des chimpanzés et s'est évanoui. On a alors vu une énorme mère chimpanzé s'approcher de lui, le prendre dans ses bras, doucement, comme s'il s'agissait de l'un des siens. Une belle démonstration de compassion... Certains animaux montrent aussi des preuves de culpabilité. On sait bien qu'un chien, par exemple, lorsqu'il a bravé un interdit, donne l'impression de se sentir coupable... Parmi les gorilles, où il y a des mâles dominants, on voit des conduites très proches de ce que nous appelons, nous, fierté ou orgueil: les dominants bombent le torse, les animaux soumis s'abaissent... Ces comportements que l'on observe chez les animaux évolués sont une esquisse de sens moral. «Quand nous ressentons une joie ou une tristesse, celle-ci est toujours en relation avec ce que nous avons vécu ou ce que nous allons vivre»
Si d'autres animaux éprouvent des sentiments, quelle est alors la spécificité de l'homme dans cette histoire ?
C'est la richesse extraordinaire qui résulte de sa grande capacité de mémoire. Nos sentiments à nous sont complexes et profonds parce que, à tout moment, notre cerveau peut se faire une idée de notre passé et de notre futur. Quand nous éprouvons une joie intense ou une profonde tristesse, celle-ci est toujours en relation avec ce que nous avons vécu ou ce que nous allons vivre. En cela, nous sommes très différents des animaux : les chimpanzés et les bonobos n'ont pas ce pouvoir de donner un sens à leur passé et à leur futur, ce qui limite forcément la profondeur de leurs sentiments. Et puis, bien sûr, il y a notre langage, qui donne une autre dimension. Avec les mots, nous pouvons faire des catégories, comparer, délibérer, choisir...
A-t-on identifié, dans nos cerveaux, des zones qui correspondraient à ces fameux sentiments ?
Grâce aux caméras à positrons, on peut visualiser les zones actives du cerveau à certains moments. On a découvert par exemple que l'amygdale (pas celle de la gorge, celle du cerveau) est liée au déclenchement de la peur et de la colère. Si vous vous trouvez nez à nez avec un ours, votre cortex visuel va envoyer des signaux à l'amygdale, qui déclenche la réaction de peur. Nous avons mené diverses expériences en demandant à des personnes sous scanner de penser à un épisode émotionnel de leur vie, et nous avons constaté que certains ensembles de neurones sont mobilisés pour certaines émotions, que les cartes cérébrales de la joie sont différentes de celles de la tristesse.
Y a-t-il une carte cérébrale de l'amour ?
On peut identifier des zones impliquées dans les processus d'excitation sexuelle et de désir, mais prétendre qu'il y aurait une région du cerveau spécifique à l'amour serait stupide. Ce sentiment met en jeu une infinité de choses. L'amour d'une personne, de la nature, ou d'une œuvre d'art, ne mobilise pas les mêmes cartes de neurones. Si vous contemplez Guernica, votre cerveau va composer une émotion esthétique et un sentiment de plaisir en effectuant la synthèse entre la vision du tableau et les idées que vous lui associez. Vous allez penser à Picasso, à la guerre d'Espagne, peut-être à des événements personnels.
Tout cela nous incite à davantage de modestie. En somme, le monde animal est un peu moins éloigné de nous que nous le croyons.
Absolument. Il nous faut admettre que l'homme n'est que le prolongement de la longue histoire de l'évolution biologique animale, et que nous avons malgré tout beaucoup de choses en commun avec les animaux. Sommes-nous supérieurs ? Bien sûr ! Nous le sommes par notre langage, notre mémoire organisée, notre mode de raisonnement complexe et notre capacité à utiliser nos sentiments pour inventer ces choses nouvelles que sont la culture et l'Histoire.
Les conséquences de ces recherches sont immenses. On pourra envisager d'intervenir sur les émotions, voire sur les comportements sociaux, ce qui peut faire aussi frémir.
Il faut certainement que la société protège ses valeurs, et qu'on ne donne pas la possibilité à une corporation de contrôler les gens. Mais, comme toujours, il y a deux faces au progrès scientifique. Mieux comprendre la nature humaine ne peut que nous aider. Mieux connaître la genèse des émotions et des sentiments nous permettra aussi d'éclairer les conflits entre les individus, de réagir plus intelligemment aux manipulations dont nous pouvons être l'objet. Les applications en bio-médecine sont nombreuses. Plus nous comprendrons le mécanisme cérébral des émotions, plus nous pourrons soulager les gens qui souffrent de dépression, syndrome qui se développe autour du sentiment de tristesse. Cela est également utile dans le traitement de la douleur: lorsque l'on souffre physiquement, on est aussi envahi par le sentiment de souffrance. On devrait pouvoir le traiter. Je crois que de telles interventions thérapeutiques seront aussi importantes que les traitements contre la polio ou le sida.
Le philosophe George Steiner nous l'avait dit ici: la culture ne nous rend pas plus humains. Mieux connaître le fonctionnement de nos sentiments ne nous rendra pas forcément plus sentimentaux ni plus civilisés.
Nous avons maintenant ouvert une petite porte pour mieux comprendre nos sentiments et nous avons la responsabilité de faire des choix. Ce que nous appelons «relations sociales» ou «culture» ne vient que de nos cerveaux. Ce sont les cerveaux qui produisent et véhiculent des comportements, des romans, des poèmes, ou des lois. D'une certaine manière, on peut voir les règles sociales et éthiques, les institutions comme des prolongements de la recherche d'équilibre que l'évolution a menée. Il est pour moi incompréhensible que certains êtres humains ne soient pas capables de ressentir de la compassion. Ce qu'ont fait les nazis est hors de mon entendement, mais, hélas ! ils ne sont pas les seuls dans l'Histoire. Cependant, les humains ne se contentent pas de montrer de la compassion comme les bonobos. Ils savent qu'ils ressentent de la compassion. Spinoza disait: «Le bonheur consiste pour l'homme à vouloir conserver son être.» Mais on peut ajouter avec lui que, si nous n'avons pas de souci pour les autres, il est impossible de continuer avec nous-mêmes.
Timing des émotions :
La variété des émotions :
L'universalité des expressions émotionnelles révèle bien que le programme émotionnel d'action n'est pas acquis et est automatisé. Chaque fois, l'émotion peut par exemple être modulée, moyennant de petits changements dans l'intensité ou la durée des mouvements qui le composent. Cependant, la routine du programme de base est stéréotypée, à tous les niveaux corporels auxquels il est exécuté — mouvements externes, modifications viscérales dans le coeur, les poumons, les entrailles et la peau, changements endocriniens. L'exécution de la même émotion peut varier selon les occasions, mais pas assez pour qu'elle ne soit plus reconnaissable par le sujet ou autrui. Elle varie autant que l'interprétation de Summertime de Gershwin peut changer selon les interprètes ou même chez le même en différentes occasions. Elle est toujours parfaitement identifiable en vertu du contour général que conserve le comportement.
Le fait que les émotions soient des programmes d'action qui ne sont pas acquis et sont automatisés et stables trahit leur origine dans la sélection naturelle et les instructions fournies par le génome qui en résulte. Ces dernières se sont conservées à travers l'évolution et se traduisent par le fait que le cerveau est assemblé d'une façon particulière de sorte que certains circuits de neurones peuvent traiter des stimuli émotionnellement compétents et inciter les régions déclenchant les émotions à élaborer des réponses émotionnelles accomplies. Les émotions, ainsi que les phénomènes qui leur sont sous-jacents, sont si essentielles pour l'entretien de la vie et la maturation subséquente de l'individu qu'elles se sont certainement déployées précocement au cours du développement.
Le fait que les émotions ne soient pas acquises, soient automatisées et soient définies par le génome éveille le spectre du déterminisme génétique. N'y a-t-il donc rien de personnel et d'appris dans nos émotions ? Bien des choses, au contraire. Le mécanisme essentiel des émotions dans un cerveau normal est assez similaire entre les individus, ce qui est un bienfait car cela procure à l'humanité, au sein de cultures diverses, un fonds commun de préférences fondamentales en matière de douleur et de plaisir. Mais il semble bien que, si ces mécanismes sont très semblables, les circonstances dans lesquelles certains stimuli sont devenus émotionnellement compétents pour le lecteur ne sont pas tout à fait les mêmes que pour moi. Il existe des choses que le lecteur craint et pas moi, et vice versa ; des choses que le lecteur aime et pas moi, et vice versa, et bien d'autres que nous redoutons l'un et l'autre. En d'autres termes, la customisation des réponses émotionnelles est considérable au regard du stimulus qui les déclenche. Influencés par la culture au sein de laquelle on nous a élevés ou par suite de notre éducation individuelle, nous avons la possibilité de contrôler en partie l'expression de nos émotions. Nous savons tous à quel point les manifestations du rire et des larmes sont différentes selon les cultures et comment elles sont façonnées, même au sein d'une même classe sociale. Les expressions émotionnelles se ressemblent, mais ne sont pas équivalentes. Elles peuvent se moduler et devenir propres à un groupe social.
Sans doute l'expression des émotions peut-elle se moduler de façon volontaire. Cependant, rien ne prouve que le degré de contrôle de ces modulations émotionnelles va au-delà des manifestations extérieures. Comme les émotions incluent bien d'autres réponses, dont plusieurs sont internes et invisibles à l' oeil nu pour autrui, le gros du programme émotionnel est encore exécuté, quelle que soit la puissance de volonté qu'on met à l'inhiber. Surtout, les sentiments émotionnels, qui résultent de la perception de tout le concert des changements émotionnels, interviennent encore même si les expressions émotionnelles extérieures sont en partie inhibées. Quand vous êtes face @? une personne stoïque qui fait à peine la moue quand on lui annonce un drame, ne croyez pas qu'elle ne ressent pas d'angoisse ni de peur. Un vieil adage portugais dit ainsi : «Voir un visage, ce n'est jamais voir le coeur.»
Gamme des émotions :
L'autre grand groupe est celui des émotions sociales. Ce label est un peu étrange, puisque toutes les émotions peuvent être sociales et le sont souvent, mais il se justifie par le cadre social impliqué dans ces phénomènes en particulier. Les exemples principaux sont la compassion, l'embarras, la honte, la culpabilité, le mépris, la jalousie, l'envie, l'orgueil et l'admiration. Ces émotions sont bien déclenchées par des situations sociales et elles jouent assurément un rôle majeur dans la vie des groupes sociaux. L'opération physiologique des émotions sociales n'est en rien différente de celle des émotions mentionnées plus haut. Elles nécessitent un stimulus émotionnellement compétent ; elles reposent sur des sites déclencheurs spécifiques ; elles sont constituées de programmes d'action élaborés qui impliquent le corps ; et elles sont perçues par le sujet sous la forme de sentiments. Cependant, on note certaines différences. La plupart des émotions sociales sont récentes en termes d'évolution et certaines seraient même exclusivement humaines. Cela semble être le cas de l'admiration et de la variété de compassion qui se polarise sur la douleur mentale et sociale plutôt que physique. Bien des espèces, et surtout les primates et les grands singes en particulier, font preuve de précurseurs de certaines émotions sociales. La compassion pour les difficultés physiques, l'embarras, l'envie et l'orgueil sont de bons exemples. Les singes capucins paraissent réagir à la perception d'injustices. Les émotions sociales impliquent un certain nombre de principes moraux et forment le fondement naturel des systèmes éthiques.
Vivre avec ses émotions (Cap Sciences)
La jeunesse des émotions
Les bébés
L’enfance
L’adolescence
A - LA COLÈRE
Un ensemble de mots ou de comportements perçus par le cortex cérébral (partie évoluée et consciente de notre cerveau) est interprété comme une menace par l'amygdale qui active l’hypothalamus, plaque tournante des émotions et grand distributeur d’hormones dans le corps.
L’adrénaline fait se dresser les cheveux, augmente la pression sanguine.
L’hypothalamus active aussi la substance grise qui orchestre les manifestations extérieures de la colère : sourcils froncés, cris, poings crispés, pli du front, gestes violents et saccadés.
La colère submerge l’affect mais il est possible de s’en accommoder et d’en faire bon usage grâce au cortex. La mémorisation des expériences passées vient enrichir notre expérience subjective. La prochaine fois nous réagirons mieux.
L’expression de la colère joue un rôle d’avertissement et permet la conciliation. Nous sommes capables d’identifier la colère dans la voix avant même de comprendre le sens de ce qui est dit. Elle révèle l’animosité d’autrui et impose d’adapter son comportement (voix douce, posture d’humilité, sourire, gestes de conciliation).
Vous dites à un homme : « Excusez-moi » et il répond avec méchanceté : « C’est quoi votre problème ? » Si vous exprimez votre colère le conflit pourrait s’envenimer. Si vous lui répondez plutôt que de réagir au problème ce sera néfaste. Si vous pouvez réévaluer la situation, imaginer qu’il a peut-être un souci aujourd’hui qui l’a rendu irritable et soupe au lait vous commenceriez à le comprendre. Si vous pouvez compatir à son souci il pourrait vous aider avec le vôtre.
L’expression de la détermination à combattre, sans qu’il y ait nécessairement de combat, a permis de découpler partiellement cette émotion du comportement d’agression.
La colère permet d’évacuer des tensions nerveuses accumulées et prévient le risque d’attaque cardiaque. Laisser sortir cette colère est salvateur, à condition de ne pas se faire mal, de ne pas faire mal à quelqu’un, de ne rien casser.
B - L'ENNUI
C - L'HUMEUR
D - L'IMPATIENCE
E - LA JOIE
F - LA NOSTALGIE
La nostalgie n’est pas propre à une culture particulière.
La nostalgie serait une méthode naturelle pour se sentir mieux, une source de bien-être psychique.
Que faut-il conseiller aux nostalgiques ? De se plonger avec émotion et regret dans la contemplation de leur passé, sans redouter d’effets néfastes. Le fait de se remémorer la couleur de sa première voiture ou son premier nounours pourrait même favoriser l’équilibre psychique. L’optimisme qui en résulte confère une bonne protection contre les idées noires.
G - LA PEUR… (voir aussi Anxiété , Stress et SIA)
La peur du noir
Focus sur le circuit de la peur : par Joseph LEDOUX (Le cerveau des émotions)
Les voies basses et hautes du thalamus vers l'amygdale :
Les neurones de l'aire thalamique qui se projettent vers le cortex auditif ou visuel primaire sont très sélectifs, et ne répondront qu'à certains signaux. En revanche, ceux de l'aire se projetant vers l'amygdale sont moins difficiles, ils seront activés par une gamme beaucoup plus large de stimuli. Les Beatles et les Rolling Stones se ressembleront pour l'amygdale vus du côté des projections thalamiques mais seront très différents vus du côté des projections corticales. Quand deux stimuli similaires sont utilisés dans un conditionnement, le thalamus envoie à l'amygdale essentiellement la même information, quel que soit le type de stimulus dont il s'occupe, alors que le cortex fera la différence entre les deux stimuli et enverra à l'amygdale des signaux distincts.
Le rôle majeur de l'amygdale :
L'axe de la peur :
L'amygdale est comme au centre d'une roue. Elle reçoit des entrées de bas niveau des régions sensorielles du thalamus, des informations de plus haut niveau du cortex sensoriel et de plus haut niveau encore (indépendants des sens) sur la situation générale provenant de l'hippocampe. Grâce à ces connexions, elle est capable de traiter la signification émotionnelle de stimuli individuels comme de situations complexes. L'amygdale est, par essence, impliquée dans l'évaluation de ce qui a un sens du point de vue émotionnel. C'est là où les stimuli déclencheurs agissent.
On peut facilement imaginer comment des dysfonctionnements de l'amygdale et de ses partenaires neuraux peuvent être à l'origine de troubles émotionnels. Si, pour des raisons génétiques ou acquises, certaines personnes ont une voie thalamique dominante ou découplée des voies corticales, elles pourraient avoir des souvenirs émotionnels sur la base de stimuli qui ne coïncideraient pas avec les perceptions conscientes courantes du monde extérieur prises en charge par le cortex. En effet, comme la voie thalamique vers l'amygdale sort du système sensoriel avant que des perceptions conscientes aient pu être créées au niveau cortical, le traitement produit par cette voie sous-corticale qui ne représente que des traits ou des fragments de stimuli ne coïncide pas forcément avec les perceptions arrivant au cortex. De telles personnes n'auraient qu'une conscience limitée de ce qui les émeut. Et si le système hippocampique était découplé des projections thalamiques et corticales vers l'amygdale, nous pourrions avoir des personnes qui expriment des émotions inappropriées au contexte immédiat, y compris social. Il s'agit pour le moment de pures spéculations, mais elles sont cohérentes avec ce que nous savons actuellement.
Le rôle de l'hippocampe :
Voies cérébrales de la défense :
EXTRACTION DES INFORMATIONS PAR LE CERVEAU
Par Jean-Philippe Lachaux :
Puis, dans un deuxième temps, l'échange d'informations entre l'avant et l'arrière du cerveau se fait selon un mode plus interactif, au cours duquel les niveaux les plus élevés de la hiérarchie semblent interroger les régions de plus bas niveau pour « approfondir l'enquête ». Cette phase d'analyse de l'information sensorielle semble pilotée depuis le sommet, ce qui justifie le nom anglais de voie top-down – du haut vers le bas – ou feedback – vers l'arrière, selon que l'on fait référence à la position hiérarchique ou anatomique.
Tout se passe comme dans les films policiers, quand un crime vient d'être commis : le poste de police prévient immédiatement le commissaire – c'est la voie bottom-up ou feedforward qui débarque ensuite toutes sirènes hurlantes sur les lieux du crime pour examiner la scène dans ses moindres détails et interroger toutes les personnes présentes – c'est la voie top-down ou feedback. Pendant la phase bottom-up, ce sont les niveaux les plus bas qui choisissent les informations dignes d'être transmises au niveau supérieur. L'agent de police prévient le commissaire parce qu'il estime qu'il s'est passé quelque chose d'important. Pendant la phase top-down, ce sont les niveaux les plus élevés qui décident de ce qui est important et de ce qui ne l'est pas.
Qu'il s'agisse d'embaucher un candidat, d'assurer le maintien de l'ordre ou d'interpréter des signaux sensoriels, le problème est finalement toujours un peu le même : trop d'informations à traiter en trop peu de temps. Le directeur des ressources humaines ne peut pas recevoir tous les candidats, le commissaire de police ne peut pas être partout à la fois et le cortex préfrontal ne peut pas réagir à la moindre stimulation sensorielle. Une fois enclenchée, chaque analyse détaillée prend du temps, beaucoup de temps. Quand le commissaire décide de se déplacer sur le terrain, il devient physiquement indisponible pour tous les autres délits. Il est donc hors de question de lui demander d'intervenir à chaque vol de vélo ou à chaque bagarre de rue. Le cortex préfrontal est soumis aux mêmes contraintes. Des phénomènes étranges, comme le clignement attentionnel que je décrirai en détail plus loin, montrent qu'une fois les régions frontales engagées il leur faut ensuite un bon tiers de seconde avant de pouvoir s'intéresser à un autre événement. Le cortex préfrontal n'est donc pas conçu pour traiter en temps réel toutes les informations provenant du monde extérieur.
Notre rapport au monde repose nécessairement sur un système d'analyse de l'information sensorielle de bas niveau, chargé de traiter en parallèle et de la façon la plus automatique possible un très grand nombre d'événements, afin de ne soumettre aux régions de haut niveau que ceux vraiment dignes d'intérêt... et revoilà le filtrage.
Cette première vague d'analyse est pré-attentive, car elle précède la sélection attentionnelle ; tous les signaux qui parviennent à nos sens doivent être pris en compte, sans a priori, de la même façon que le poste de police est censé consigner tous les délits qui lui sont rapportés. Seuls certains cas font ensuite l'objet d'une enquête approfondie. Cette phase d'analyse repose sur les neurones détecteurs que nous avons déjà évoqués. Ces neurones analysent à tout moment notre environnement de façon rapide, automatique et massivement parallèle, pour y détecter la présence éventuelle de caractéristiques particulièrement intéressantes : ici du bleu, là un objet se déplaçant vers la droite et peut-être là-bas un visage ou un animal. En un clin d'oeil, l'information se propage selon le mode feedforward jusqu'au cortex préfrontal où elle déclenche, ou non, une seconde phase d'analyse plus détaillée selon le mode feedback. Si c'est le cas, le visage ou l'animal est identifié de façon précise et le cerveau peut décider de la conduite à tenir. Le stimulus a attiré l'attention.
2 - Critères de décision :
Il ressort de ces observations que le processus de filtrage prend toujours en compte plusieurs critères, dont les plus simples, comme ici la couleur, ne suffisent pas toujours à éliminer tous les distracteurs. Il n'est pas impossible que le cortex préfrontal intervienne lui-même dans ce type de vérification complexe, très rapidement, et avant de décider de s'engager pleinement ou non. Certains experts trouvent d'ailleurs le terme « filtrage attentionnel » un peu exagéré et préfèrent parler d'un phénomène d'atténuation, car un vrai filtre ne laisserait passer aucun des mots rouges. Que diriez-vous d'un filtre à café qui laisserait passer des petits grains au fond de la cafetière ? Dans le cas des histoires, il s'agit effectivement plutôt d'un processus d'atténuation de la vague d'activité neuronale générée par les mots à ignorer. Mais la plupart des systèmes de filtrage de la vie courante fonctionnent aussi comme des atténuateurs plus que comme des filtres à café. Au bout d'un certain temps, quelqu'un ou quelque chose parvient toujours à tromper la vigilance du système et à s'infiltrer. Prenez le cas du dispositif de sécurité dans les aéroports : bien qu'il s'agisse indéniablement d'une forme de filtre, des passagers parviennent régulièrement à le déjouer pour s'infiltrer à bord des avions munis d'objets dangereux. Le dispositif n'élimine pas le danger, mais il l'atténue.
Mais pourquoi certaines informations sensorielles déclenchent-elles une réaction des régions frontales et d'autres pas ? En songeant à cette question, je ne peux m'empêcher de penser au courageux Philippidès qui lors de la bataille de Marathon, courut 42 kilomètres pour annoncer la victoire d'Athènes avant de mourir d'épuisement. Comment prévenir l'avant du cerveau quand on est perdu tout à l'arrière du lobe occipital ? À première vue, il est d'abord essentiel que la vague d'activité générée par le stimulus puisse se propager jusqu'au lobe frontal sans perdre de sa force pendant la phase ascendante feedforward. Pour cela, les neurones disposent de plusieurs astuces, de bon sens, qui sont celles qui nous viendraient naturellement à l'esprit dans une situation similaire. Comme nous ne sommes pas des neurones, je vous propose d'imaginer une situation plus humaine proche de celle à laquelle sont confrontées les cellules du cortex : un groupe de joueurs de percussions est échoué sur une île absolument déserte et cherche à se faire remarquer d'un navire passant au loin, sans rien d'autre sur cette île que leurs instruments échoués avec eux. Chaque joueur représente un neurone, qui déclenche un potentiel d'action à chaque coup de tambour, et le bateau représente le lobe frontal, ou une région du cortex située plus haut dans la hiérarchie du système visuel, qui pourrait à son tour prévenir le lobe frontal. L'un des musiciens aperçoit le bateau depuis la plage. Par réflexe professionnel, il s'empare d'un tambour et tape dessus, dans l'espoir naturel d'être entendu du navire. Mais plusieurs jours sans manger ont eu raison de sa vigueur : il frappe un coup, puis s'arrête, épuisé, puis recommence et s'arrête à nouveau ; le son est si faible que l'équipage n'entend rien. Il faudrait jouer plus fort, beaucoup plus fort, et plus vite, mais le pauvre homme en est incapable. Il part donc chercher du renfort. Bientôt, les cent naufragés de l'île tapent sur leur tambour dans une sorte de cacophonie faiblarde : tandis que certains jouent, d'autres reprennent leur souffle, si bien qu'au total, il n'y a jamais plus de dix tambours qui résonnent en même temps. « Il faudrait être dix fois plus nombreux ! », dit l'un d'eux, mais tout le monde est déjà sur la plage. Un autre réalise tout d'un coup leur erreur : « Nous devons tous frapper en même temps ! » Mais bien sûr ! Les naufragés décident aussitôt de synchroniser leurs efforts pour jouer à l'unisson. Le son est cette fois si fort qu'il parvient aux oreilles d'un marin, qui déroute le bateau en direction de l'île pour sauver les naufragés. Moralité : pour se faire entendre, il faut soit être fort, soit être nombreux et organisé — soit les deux, évidemment.
Ce qui est vrai sur une île déserte l'est aussi dans le cerveau. Les neurones utilisent ce genre d'astuces pour se faire entendre des autres régions cérébrales, comme l'ont montré les Américains John Reynolds et Bob Desimone en étudiant l'aire visuelle V4. V4 est l'une des régions du cortex visuel (d'où la lettre V) chargées d'analyser, à un niveau assez simple, les images qui viennent chatouiller la rétine. V4 est située un peu plus haut que V1 dans la hiérarchie du système visuel, ce qui lui vaut son numéro supérieur, et ses neurones s'y intéressent à des aspects de l'image un peu plus sophistiqués que dans V1, comme la présence de certaines formes particulières que les neurones de V1 ou V2 sont incapables de reconnaître. Le travail de Reynolds et Desimone a consisté à enregistrer la réponse des neurones de V4 à des stimuli visuels dans le cortex du singe, pour étudier comment cette réponse varie en fonction de l'attention. L'idée d'aller voir dans V4 était particulièrement astucieuse, non seulement parce qu'il est relativement facile d'y insérer des électrodes, mais aussi parce que les neurones y sont déjà spécialisés dans la détection de caractéristiques assez intéressantes, comme la forme et la couleur, tout en ayant des champs récepteurs bien définis. Comme dans la plupart des aires visuelles, notamment celles dites primaires, chaque neurone de V4 a effectivement un champ récepteur, c'est-à-dire une région du champ visuel qu'il a charge de surveiller. Avec beaucoup de patience, il est possible d'entraîner un singe à porter son attention vers n'importe quel endroit du champ visuel, et en particulier vers celui que le neurone surveille. Il suffit alors de présenter un stimulus visuel dans ce champ récepteur pour étudier comment varie la réaction du neurone en fonction de la direction de l'attention : selon qu'elle est orientée vers ce champ récepteur ou ailleurs.
C'est exactement ce que firent Reynolds et Desimone, qui constatèrent que l'effet de l'attention dépend fortement du contraste du stimulus montré à l'écran. Je rappelle que le contraste correspond à peu près à la différence de luminosité entre le stimulus et le fond sur lequel il apparaît. Une forme gris clair présentée sur un fond à peine plus clair a un contraste faible, comme un objet émergeant à peine du brouillard. Une fois le brouillard dissipé, le même objet au même endroit a un contraste élevé, il est plus visible. Vous savez bien que, les jours de brouillard, il faut faire plus attention en conduisant. Vous allez maintenant savoir aussi pourquoi.
Les résultats de Reynolds et Desimone peuvent se résumer ainsi : lorsqu'on présente à un neurone, dans son champ récepteur, une forme au contraste si faible qu'elle en est invisible, ce neurone ne réagit pas, exactement comme si la forme n'était pas là. La forme est pourtant présente à l'écran, mais elle est très en dessous du seuil de perception ; et, dans ce cas, l'attention ne peut rien pour elle : même si l'animal a l'attention rivée sur la forme, le neurone ne réagit pas. Si le contraste est au contraire élevé, et que la forme est bien visible, ce même neurone réagit en produisant une belle bouffée de potentiels d'action, mais cette réponse n'augmente pas non plus avec l'attention : qu'il fasse attention à la forme ou ailleurs, la réponse du neurone est la même. La forme étant de toute façon visible, elle ne devient pas plus visible sous l'effet de l'attention. Ce n'est qu'à des niveaux de contraste intermédiaires que l'effet de l'attention se fait sentir, lorsque la forme est à peine visible, juste au-dessous, ou juste au-dessus, du seuil de perception. Dans ce cas, le neurone répond au stimulus avec beaucoup plus de vigueur si l'attention est dirigée vers la forme ; le nombre de potentiels d'action augmente comme si le contraste du stimulus était plus élevé, et l'animal réagit effectivement comme s'il voyait mieux la forme présentée. Dans V4, et ailleurs, jusque dans les régions sous-corticales, l'attention a donc pour effet d'augmenter la visibilité des objets difficiles à discerner. C'est pourquoi il faut être plus attentif quand on conduit dans le brouillard ou la nuit. Dans ces conditions, l'attention permet d'y voir plus clair et donc de réagir plus vite.
Cet effet permet à l'activité neuronale de V4 de mieux se propager en direction du lobe frontal. N'oublions pas qu'un peu plus haut dans la hiérarchie du système visuel, d'autres neurones sont à l'écoute de leurs collègues de V4 qui doivent leur signaler ce qu'ils voient en envoyant des potentiels d'action le long de leurs axones, un peu comme les amis de l'aveugle doivent lui signaler ce qu'ils voient pour l'aider à traverser la rue. Quand les neurones de V4 chargés de surveiller une région du champ visuel restent silencieux, les neurones situés à l'étage au-dessus en déduisent qu'il ne se passe rien et que la région est vide. La transmission de l'information s'arrête là et n'atteindra donc jamais le sommet de la hiérarchie. L'intérêt de ce mécanisme d'amplification de l'activité neuronale quand le stimulus est à peine visible est donc évident : lorsque vous portez votre attention quelque part autour de vous, les neurones de V4 qui ont en charge cette région réagissent exactement comme si le contraste de l'image y était plus marqué, comme si le brouillard était moins épais à cet endroit. Ils rompent leur silence et se mettent à signaler la présence d'une forme dans leur champ récepteur, par exemple celle d'un panneau de signalisation à peine visible dans la brume. Cette information est ensuite relayée vers le sommet de la hiérarchie qui réagira pour mieux étudier ce panneau.
Ce mécanisme est si efficace que, lorsque l'attention se porte sur une région de l'espace, les neurones dont le champ récepteur occupe cette région deviennent hypersensibles, au point de s'activer avant même qu'un stimulus n'y apparaisse : ils se préparent à voir quelque chose. C'est un phénomène connu en anglais sous le nom de baseline shift, c'est-à-dire, une augmentation du niveau d'activité de base. Les neurones grésillent d'impatience, avec une augmentation d'activité qui peut atteindre 30 %. Un murmure grandissant envahit les tribunes avant l'explosion de joie au moment du but.
Ce mécanisme de « préchauffe » contribue à amplifier la réponse neuronale à l'apparition d'un stimulus et amène ainsi le sujet à réagir plus vite, comme dans l'expérience de Posner. Il s'agit d'ailleurs d'un phénomène général, observé non seulement dans V4, mais également dans de nombreuses autres régions du cerveau. Selon que le cerveau va devoir faire attention à la couleur ou au mouvement d'un objet par exemple, le murmure des neurones augmente dans l'une ou l'autre des régions spécialisées dans l'analyse de ces deux aspects, avant même que l'objet n'apparaisse.
Formidable, n'est-ce pas ? Il suffit donc qu'au moment de chercher ma raquette de tennis dans le placard, je « préchauffe » mes neurones spécialisés dans la détection de ma raquette, pour la trouver immédiatement. Hum... il y a quand même quelque chose qui cloche. Réfléchissons une seconde : si c'était le cas, pourquoi M. Couleur ne trouve-t-il pas immédiatement la paire de gants noirs dans le tiroir ? En réalité, ce système de préchauffe a deux limites. La première, que nous avions déjà évoquée, c'est qu'il n'existe pas forcément dans mon cerveau de neurones spécialisés dans la détection des gants noirs ou des raquettes de tennis. La seconde limite, c'est que même si de tels neurones existaient, il faudrait encore que le cerveau sache les trouver pour les préparer à voir la raquette. Cela peut paraître bizarre, j'en conviens ; le cerveau ne saurait pas où sont ses neurones ? Tout cela demande quelques explications.
Vous admettrez sans peine que cet effet de préchauffe a forcément son origine dans le cerveau. Quand l'attention se porte sur une zone particulière du champ visuel, c'est parce que certaines régions du cerveau ont réussi à pré-activer sélectivement les neurones de V4 chargés de surveiller cette zone. Ces régions, dont je parlerai en détail plus tard, sont situées dans le lobe pariétal et dans le lobe frontal. Elles peuvent modifier l'activité neuronale dans V4 parce qu'elles y ont accès, grâce à des connexions à longue distance relativement fixes. Ces connexions leur permettent de « joindre » les neurones de V4, comme on joindrait un correspondant au téléphone. S'il leur est si facile de moduler l'activité de tous les neurones chargés de surveiller la même zone du champ visuel, c'est parce que ces neurones se trouvent les uns à côté des autres. Si les neurones chargés de surveiller la zone en question étaient placés n'importe comment, les régions frontales et pariétales souhaitant les pré-activer auraient beaucoup plus de mal à les aider de façon sélective, sans faciliter également tous les autres neurones. De façon générale, le mécanisme de préchauffe attentionnel fonctionne donc bien pour des groupes de neurones proches les uns des autres, pour des raisons simples de connectivité. Si vous deviez prévenir demain les 260 000 habitants de Strasbourg qu'ils doivent se préparer à vivre un événement exceptionnel, vous pourriez y arriver, moyennant un certain budget, par le biais d'affiches et d'annonces dans les journaux locaux. Mais vous auriez plus de mal à prévenir chacun des 240 000 Français portant le nom de Martin, parce qu'ils se répartissent sur tout le territoire. Dans le cerveau, la détection de certaines propriétés repose parfois sur des neurones proches les uns des autres, comme c'est le cas pour la position dans l'espace, la direction du mouvement ou la couleur. Ces neurones sont plus faciles à pré-activer et les propriétés correspondantes sont plus facilement la cible de l'attention.
Heureusement, tout cela n'est pas gravé dans le marbre. Il est toujours possible, à force d'apprentissage, de se sensibiliser à d'autres aspects du monde. Revenons à notre enquête policière : si le commissaire pense que le criminel rôde dans le XIII' arrondissement de Paris, il peut facilement demander aux policiers qui y sont en faction de redoubler de vigilance et de lui signaler le moindre événement anormal. Il porte alors son attention sur une région de l'espace, comme le ferait le lobe frontal en agissant sur V4. S'il sait que le criminel fréquente le milieu des bars, il peut demander aux services de police spécialisés dans ce type de commerce d'être plus vigilants. Il porte alors son attention sur une propriété simple, comme le cerveau porterait attention à la couleur bleue. Tout cela est facile. Mais si le criminel semble se plaire dans les bars possédant le jeu vidéo Space Invaders, un Baby-foot et des fenêtres donnant au sud, la tâche du commissaire devient plus ardue, car il n'existe pas de brigade spécialisée dans la surveillance de ce type d'établissement. Il faudra donc beaucoup de temps pour trouver le malfaiteur, puisqu'il faudra inspecter tous les bars un par un. Mais si, pour une raison inconnue, de nombreux criminels se mettaient à fréquenter les bars avec jeu vidéo, Baby-foot et exposés au sud, la police finirait par créer une brigade spécialisée, et le moindre événement louche qui s'y produirait serait rapidement signalé. En neurobiologie, ce type de réorganisation porte le nom de plasticité. Lorsque certains stimuli prennent une importance particulière et durable, le cerveau est capable de « former » une brigade de neurones dans la détection de ces stimuli. Nous en avons déjà vu un exemple avec les travaux de Jonathan Fritz sur le cortex auditif du furet. Plus un stimulus est important pour le cerveau, plus l'effectif de la brigade neuronale chargée de le détecter est grand et plus ce stimulus aura du mal à passer inaperçu.
Le système de facilitation de Reynolds et Desimone peut donc théoriquement s'appliquer à la détection de propriétés assez complexes, à condition sans doute que les neurones concernés puissent être facilement « joignables », et si possible situés les uns à côté des autres. Si vous avez déjà été à la pêche au crabe, vous avez peut-être remarqué qu'après un long moment passé à chercher sans trouver, tout ce qui ressemble de près ou de loin à un crabe finit par attirer l'attention, même un petit bout d'algue noir coincé sous un rocher. Le cerveau développe une forme d'hypersensibilité à tout ce qui ressemble à un crabe. Cette hypersensibilisation peut s'expliquer par un mécanisme semblable à celui observé par Reynolds et Desimone dans V4, qui favoriserait des neurones assez sophistiqués chargés de détecter les formes caractéristiques des crabes. Selon le principe décrit par Desimone, ces neurones deviendraient alors plus sensibles et plus réactifs. Mais l'exemple de l'algue montre bien les limites de ce système : seule une deuxième phase d'analyse visuelle approfondie permet de vérifier si la forme en question est bien un crabe ou non.
Depuis quelques années, un autre mécanisme de facilitation attentionnelle semble avoir été découvert. Souvenez-vous de l'astuce trouvée par les naufragés pour se faire remarquer du navire : synchroniser leurs efforts pour mieux se faire entendre. Dans une étude menée en 2001 avec Pascal Fries, Desimone et Reynolds se demandèrent si les neurones de V4 savaient aussi utiliser la synchronisation pour augmenter leur force de frappe sur le reste du cerveau. Comme chaque portion de l'espace visuel est surveillée par de nombreux neurones de V4 ayant tous le même champ récepteur, il suffirait effectivement, en théorie, que ces neurones envoient leurs potentiels d'action en même temps pour mieux se faire entendre par ceux des niveaux supérieurs. Il restait à faire la preuve expérimentale de ce mécanisme, et c'est ce que réussit l'équipe de Desimone : les neurones de V4 ont effectivement tendance à émettre leurs potentiels d'action de façon synchrone quand l'attention est orientée sur la partie du champ visuel qui les concerne. Ils ne le font d'ailleurs pas à n'importe quel rythme, mais dans une gamme de fréquence assez large et rapide, appelée bande gamma, au-delà de 40 Hz ; je vous avais prévenu : les oscillations, c'est important pour l'attention. Sous l'influence, justement, de l'attention, les neurones s'activent de façon rythmique et en phase, comme une foule applaudissant en rythme, plusieurs dizaines de fois par seconde, parfois jusqu'à cent fois. Au contraire, si l'attention se déplace hors de cette zone, les potentiels d'action sont produits de façon plus anarchique, sans organisation temporelle précise ; chacun applaudit à son rythme, sans se caler avec ses voisins et, forcément, le son est moins fort. Le recours à la synchronisation permet donc également de renforcer l'impact d'un groupe de neurones, sans forcément passer par une augmentation du nombre de leurs potentiels d'action. Il s'agit donc d'une forme de facilitation attentionnelle « économique », comme ont pu le remarquer les musiciens sur l'île. Rien de nouveau sous le soleil, dit-on, même sous celui caressant des plages du Pacifique. Les astuces trouvées par ces naufragés sont vieilles comme le système nerveux.
Mais à quoi sert donc l'attention quand il fait jour et qu'il n'y a pas de brouillard ? Quand un neurone de V4 a dans son champ récepteur un objet bien visible, il réagit, nous l'avons vu, et prévient les neurones qui l'écoutent de la présence de l'objet. Le mécanisme d'amplification décrit par Reynolds et Desimone ne sert donc en principe à rien, et seule la synchronie pourrait sembler éventuellement utile. Mais V4 rencontre alors un autre problème que doit aussi résoudre l'attention : faire face à la complexité du monde visuel.
Prenons tout de suite un exemple concret : pour un joueur de tennis professionnel s'apprêtant à retourner le service de son adversaire, certains détails dans le geste de préparation de ce dernier, par exemple la façon dont il tient sa raquette, permettent de prédire si le service va être à plat au centre, ou slicé vers le couloir de double. Il est donc important pour le retourneur d'arriver à extraire cette information visuelle, à une vingtaine de mètres de distance. Ce travail revient en partie aux neurones de V4 chargés d'analyser la région de l'espace visuel où se trouve la main du serveur. Leur tâche est compliquée, car ils ne voient pas que la main, mais également le manche rose fluo de la raquette, les arbres derrière le court, la montre du joueur et quantité d'autres éléments inutiles et distrayants occupant leur champ récepteur. Le retourneur doit en faire abstraction pour se concentrer sur la seule information qui lui soit utile : la prise de raquette de son adversaire. Les neurones de V4 doivent donc traiter l'image exactement comme si les éléments distracteurs n'étaient pas là, en les ignorant totalement grâce à l'attention. C'est exactement ce qui se passe, grâce à un mécanisme de « compétition biaisée », selon le terme inventé par John Duncan et Bob Desimone.
Dans des conditions de bonne visibilité, le problème n'est donc pas de voir les objets, mais d'arriver à les distinguer les uns des autres. De jour et par temps clair, le monde visuel est si riche et complexe que chaque neurone de V4 ne voit pas une forme, mais plusieurs, car même s'il ne voit le monde que par un trou de serrure, la région couverte par son champ récepteur est généralement occupée par plusieurs objets à la fois. Pour comprendre comment réagit ce neurone, il faut ensuite savoir que dans V4, comme dans V1, les neurones ne répondent pas exactement avec le même enthousiasme, la même force, à tout ce qui leur est montré ; ils ont leurs petites préférences. Le champ récepteur est déjà une forme de préférence, pour les formes apparaissant à un endroit bien précis du champ visuel, mais la réaction des neurones dépend aussi de ce qui leur est présenté : certains neurones préfèrent le bleu et d'autres le rose ; certains préfèrent certaines formes simples particulières, etc. Concrètement, quand un neurone de V4 voit apparaître dans son champ récepteur son stimulus préféré – par exemple, le manche rose fluo de la raquette –, il réagit vivement en émettant une volée de potentiels d'action qui sont comme des coups de tambour prévenant les neurones situés plus haut dans la hiérarchie. Quand ce même neurone voit un stimulus qu'il apprécie moins — par exemple, la main du serveur —, il réagit plus mollement, avec par exemple deux potentiels d'action, contre dix pour son stimulus préféré. Mais si ce neurone voit ces deux stimuli ensemble, il ne réagit pas avec douze coups de tambour, comme on pourrait s'y attendre, mais avec un enthousiasme mitigé, en frappant seulement six coups. Lorsqu'un neurone de V4 voit se présenter dans son champ récepteur plusieurs formes, sa réponse est généralement une sorte de moyenne entre les réponses qui seraient les siennes si chacune des formes était présentée seule.
Lorsqu'on présente à un neurone de V4 deux figures dans son champ récepteur, sa réponse se situe à un niveau intermédiaire entre celles que l'on pourrait mesurer si chacune de ces figures était montrée seule (ce qu'illustrent Les trois écrans du haut). Toutefois, cette règle change si l'animal porte son attention à l'intérieur du champ récepteur du neurone. Dans ce cas, la réponse observée correspond à celle qu'entrainerait la présentation de la figure à laquelle fait attention le singe, si elle était montrée seule. L'attention « efface » l'autre figure.
Desimone a étudié la réaction des neurones de V4 confronté à deux objets, selon que l'attention se porte sur l'un ou sur l'autre, exactement comme si le retourneur faisait attention au manche rose fluo ou à la main du serveur. Dans ce cas, un neurone de V4 réagit presque comme si le stimulus cible de l'attention était tout seul. Si c'est à son stimulus préféré que l'animal fait attention, le neurone frappe neuf fois sur son tambour, au lieu de six. Mais si l'attention va sur l'autre stimulus, le neurone frappe trois coups. Pour un neurone situé dans la pièce d'à côté, c'est-à-dire un peu plus haut dans la hiérarchie du système visuel, tout porte à croire que V4 n'a vu qu'un seul des deux stimuli, comme si l'autre avait été gommé de l'image. L'attention agit donc en faisant croire aux neurones d'à côté que V4 n'a vu qu'un seul stimulus. C'est ce que Duncan et Desimone désignent sous le nom de « compétition biaisée ». Quand l'attention est occupée ailleurs, les deux stimuli se livrent une lutte acharnée pour contrôler l'activité du neurone. Le premier stimulus l'encourage à frapper dix coups pour montrer qu'il est là, mais le deuxième stimulus l'en empêche, car il souhaite que le neurone tape deux fois pour signaler que c'est lui qui est là. Cette bagarre sans merci aboutit à une forme de compromis qui ne satisfait personne, six coups de tambour. C'est alors qu'arrive l'attention, tel l'arbitre d'un match de tennis, pour régler la dispute en laissant l'un des deux stimuli décider. Celui qui n'a pas ses bonnes grâces n'a plus qu'à ravaler sa fierté, le point est accordé au stimulus « main du serveur ». La compétition est biaisée, c'est de la triche !
Ce mécanisme de compétition biaisée a pour seul inconvénient d'être un peu lent. Les expériences de Desimone montrent que l'arbitre-attention met deux dixièmes de seconde pour arriver jusqu'à V4 — le temps peut-être de traverser le terrain depuis le lobe frontal, au contraire de l'autre mécanisme, le baseline shift, qui permet, lui, de préchauffer les neurones avant même que le stimulus n'apparaisse, mais sans permettre de sélectionner entre la main et le manche. Si l'on enregistrait la réponse d'un neurone de V4 chez ce joueur de tennis à partir du moment où son regard se porte sur la main du serveur, on constaterait que ce neurone commence par tout « voir » le manche fluo, la main, la montre et les arbres derrière, avec cette espèce de réponse intermédiaire que nous avons mentionnée. Il faudrait attendre un cinquième de seconde environ pour que sa réponse se stabilise au niveau qui serait la sienne si le manche, la montre et les arbres avaient été effacés. Le mécanisme de compétition biaisée ne semble donc pas pouvoir fonctionner dès la première vague feedforward pré-attentive, ce qui est logique puisque cette compétition n'est résolue que par l'attention, il s'agit donc plutôt d'un mécanisme de type feedback, ou descendant. Ce retard laisse effectivement penser que l'effet se propage depuis le sommet de la hiérarchie, peut-être depuis le lobe frontal, comme une sorte de cascade du haut vers le bas, top-down. Il a d'ailleurs été montré que la compétition biaisée n'existe pas que dans V4, mais pratiquement à tous les niveaux de hiérarchie du système visuel, avec un temps de retard – par rapport à l'apparition des stimuli visuels – qui est plus court pour les zones les plus élevées dans cette hiérarchie, comme si l'arbitre venait effectivement de tout là-haut. Le biais en faveur de la main du serveur s'observe par exemple dans V4 avant de s'observer dans Vl. Le mécanisme de compétition biaisée n'est donc pas à l'origine du fait que certains stimuli allument le cortex préfrontal et d'autres non, il en est plutôt la conséquence. La capacité à activer le lobe frontal semble plutôt favorisée par le système de préchauffe, le baseline-shift, ou par la synchronie. La compétition biaisée interviendrait alors dans une deuxième phase, apparemment sous l'impulsion des régions frontales, pour affiner l'analyse d'une information sensorielle jugée potentiellement importante. Il serait donc le fruit du dialogue entre le commissaire de police et les agents sur le terrain.
Tout ceci rappelle étrangement le comportement de la vague rouge dans l'expérience des histoires. Souvenez-vous que, pendant le premier quart de seconde, la réponse du cortex visuel était la même pour les mots rouges et verts, jusqu'à ce que la vague rouge finisse par retomber. Un quart de seconde, c'est donc le temps nécessaire pour que le cerveau gomme toute trace des mots rouges – et toute trace du poignet rose fluo ; au-delà, la sélection a eu lieu, et le monde redevient enfin simple.
Ce mécanisme de compétition biaisée s'exprime à tous les niveaux dans le cerveau. Les expériences de neuro-imagerie réalisées chez l'homme montrent, de façon générale, que, dans le cerveau d'un sujet engagé dans une tâche, l'activité des régions impliquées dans le traitement des informations qui sont importantes pour la tâche augmente, tandis que celle des autres régions diminue, et ce phénomène s'amplifie avec la difficulté de la tâche. Dans les cas extrêmes où la tâche à réaliser est très difficile, l'activité de ces régions « inutiles » peut même aller jusqu'à s'annuler, c'est-à-dire jusqu'à atteindre le niveau qui serait le leur si elles n'étaient pas stimulées : l'aire visuelle V5, par exemple, est d'ordinaire active dès que le cerveau perçoit des objets qui se déplacent, mais chez une personne occupée à examiner de façon très attentive la couleur d'un objet en mouvement, l'activité de V5 est plus faible que si son attention se portait sur le mouvement. Et si l'exercice est difficile et demande une très grande attention, l'activité de V5 peut franchement s'annuler comme si l'objet ne bougeait pas. Il s'agit toujours du même phénomène de « gommage », de compétition biaisée, appliqué cette fois à une propriété — le mouvement. Et comme l'a clairement montré la chercheuse anglaise Nilli Lavie à travers plusieurs études, il est d'autant plus efficace que la tâche à réaliser demande d'être attentif.
Il n'est pas exclu que l'attention ait d'autres tours dans son sac, mais nous voyons déjà sur plusieurs exemples que le cerveau peut se débrouiller avec quelques mécanismes simples. Il faut d'ailleurs insister avant de clore ce chapitre sur le fait que les mécanismes attentionnels qui viennent d'être décrits dans le cas de la vision s'appliquent aussi aux autres systèmes sensoriels. Il y a bien sûr quelques différences dues à la différence de nature entre les images, les sons ou les odeurs, mais les principes restent à peu près les mêmes. Quand quelqu'un vous adresse la parole, par exemple, l'activité des régions du cortex auditif et du lobe frontal gauche chargées d'analyser sa voix est très diminuée si vous êtes en train de lire au lieu de l'écouter. Si vous donnez l'impression d'être « dans votre bulle », c'est donc à cause de cet effet attentionnel qui limite le traitement de toutes les stimulations extérieures qui ne concernent pas votre lecture. Et si vous relâchez un instant votre attention visuelle pour écouter ce qu'on vous dit, ce sont les zones de la lecture qui baisseront leur activité. La réponse de votre cerveau à chaque mot s'arrêtera aux portes du lobe frontal... – encore cette vague rouge. L'alternance entre des phases d'écoute et des phases de concentration sur la lecture se traduit donc par une bascule entre deux réseaux, l'un traitant la parole et l'autre chargé de la lecture, à tel point qu'une mesure en direct de votre activité cérébrale suffirait à savoir si vous écoutez ou non ce qui vous est dit. Il ne reste plus qu'à imaginer une cocktail party où chaque convive saurait enfin si ces voisins l'écoutent, grâce à de charmants dispositifs d'IRMf miniatures équipant la tête de chacun. Quelle belle soirée en perspective !
MALADIE D'ALZHEIMER
Il est toujours difficile de distinguer les premiers stades d’une démence du vieillissement normal. La difficulté tient au fait que les premiers symptômes des démences apparaissent très graduellement, souvent à l’insu de la personne atteinte.
Mais c’est une chose d’oublier un nom ou un rendez-vous de temps en temps, puis de s’en souvenir par la suite, et c’en est une autre lorsque ces oublis sont fréquents, que la personne ne s’en souvient pas plus tard, et qu’ils se mettent à nuire aux activités quotidiennes.
Avec l’évolution de la forme modérée vers la forme avancée, il n’est pas rare de voir apparaître des problèmes psychologiques plus sévères et handicapants, comme de l’insomnie, des fluctuations de l’appétit ou des changements de la personnalité qui s’ajoutent au vécu déjà difficile du patient.
Quand la démence de type Alzheimer atteint le stade avancé, la majeure partie du cortex est sérieusement endommagée. Les individus ont des difficultés graves à s’exprimer, à reconnaître leurs amis et même leurs proches. La confusion mentale, aggravée par une perte de mémoire sévère, donne souvent lieu à des illusions, des hallucinations ou des délires paranoïdes.
Pour les professionnels de la santé, les termes «léger, modéré et avancé» désignant les trois grands stades de la démence de type Alzheimer manquent un peu de précision. C’est pourquoi ils utilisent une terminologie plus clinique suivant des critères comportementaux, l'échelle de détérioration globale (EDG) ou échelle de Reisberg (d’après son auteur, le Dr. Barry Reisberg).
Échelle de Reisberg
Stade 2 : Troubles de mémoire
Stade 3 : Confusion bénigne
Stade 4 : Confusion avancée
Stade 5 : Problèmes bénins d'ordre cognitif
Stade 6 : Problèmes moyens d'ordre cognitif -- supervision constante
Stade 7 : Problèmes avancé d'ordre cognitif
L’espérance de vie pour les personnes atteintes d’une démence de type Alzheimer est en moyenne de 8 à 12 ans. La progression de la maladie varie d’une personne à l’autre et dépend en partie de l’âge du patient au moment du diagnostic et de son état de santé général.
Le nombre de personnes atteintes de la maladie d'Alzheimer augmente régulièrement : 11 millions de cas en 1980, 18 millions en 2000, 25 millions en 2004.
en France, selon l’étude Paquid de 2001, 17,8% de la population âgée de + de 75 ans soit 900 milles malades avec 220 milles nouveaux cas par an et la perspective de 1,2 million de malades en 2020 et de 2 millions vers 2040.
Alain Lieury : L'environnement enrichi pourrait même avoir des effets de préservation dans la maladie d'Alzheimer. En effet, certaines souris transgéniques développent une anomalie caractéristique de la maladie d'Alzheimer (en américain AD = Alzheimer Desease) : la fabrication de plaques à cause d'un gène mutant de la protéine A4-amyloïde qui s'accumule ; ces plaques bouchent les transmissions et envahissent le cerveau. Une équipe de neuf chercheurs américains, sous la conduite de David Costa du Centre de Recherche sur la maladie d'Alzheimer en Floride, a montré dans une expérience très complète, que non seulement l'élevage en milieu enrichi peut avoir des effets positifs sur les capacités cognitives mais que les tests d'apprentissage eux-mêmes sont bénéfiques.
Test de Gröber et Buschke utilisé pour déceler la maladie : Ce test est destiné à explorer la mémoire. Ce test présente l’avantage de limiter l’interférence des troubles de l’attention et de compréhension. Il va permettre de comparer les capacités d’encodage et de consolidation et l’efficacité des mécanismes de récupération.
Dans ce test le patient sera aidé:
Anatomo-pathologie : Le cerveau du patient est victime d'un double processus de dégénérescence et d’inflammation, caractérisé par deux types de lésions locales participant à une lésion plus globale du cerveau :
MALADIES OU TROUBLES NEUROPSYCHIQUES
Denis le Bihan : le docteur Insel, directeur de l'institut national de la santé mentale au NIH a déclaré qu'en 2012 les pathologies psychiatriques devraient être redéfinies sur la base d'anomalies des circuits cérébraux, et d'interconnexions entre régions cérébrales...
Par Sylvain Bartolami : Les maladies du système nerveux central se divisent en deux grandes catégories : les maladies associées à des anomalies ou des lésions du tissus nerveux et les maladies dépourvues de signes somatiques ou moléculaires. Les premières sont les maladies neurologiques, les secondes les maladies psychiatriques. Cette classification simpliste induit nécessairement un flou dans la frontière entre ces 2 grandes familles de pathologie, d’autant plus que cette classification est en évolution permanente selon les avancées des recherches en neurobiologie. Une maladie psychiatrique pouvant changer de catégorie avec la découverte d’anomalies cellulaires ou géniques expliquant au moins en partie sa physiopathologie. Inversement une maladie neurologique peut développer des complications d’ordre psychiatrique. Aussi, cette classification « académique » peut être remise en question à tout instant. Elle doit être abordée avec quelques précautions et sans à priori dogmatique. Une autre manière de subdiviser les maladies du SNC serait de les définir sur une base psycho-cognitive plutôt que anatomo-pathologique. Dans ce cadre, lorsque la pensée et/ou l’humeur du malade est directement affecté alors la pathologie concernera la sphère psychiatrique, qu’il y ait ou non des lésions anatomiques et/ou des anomalies génétiques impliquées dans la physiopathologie. Quoiqu’il en soit ces troubles neuropsychiatriques sont un important problème de santé publique comme l’attestent certains chiffres : 70 millions de personnes en souffrent aux USA et plus d’un million de français sont soignés chaque année pour de tels troubles.
Agraphie et Alexie :
Incapacité d'écrire et de lire. Ces deux troubles du langage semblent dépendre de deux systèmes spécialisés distincts et autonomes.
Alexithymie : terme introduit en 1970 par John Nemiah et Peter Sifneos. L'alexithymie (du grec a : préfixe privatif, lexis signifiant « mot » et thymos signifiant « humeur ») désigne les difficultés dans l’expression verbale des émotions communément observées parmi les patients présentant des symptômes psychosomatiques.
Selon les observations de Nemiah et Sifneos, les patients psychosomatiques ont en commun une difficulté marquée à décrire leurs sentiments, une absence ou une réduction de la vie fantasmatique, et la manifestation de la pensée opératoire. À cette définition, sont venus se greffer des éléments descriptifs discutables, comme : personne ennuyeuse, terne, morne, au fort conformisme social, voire incapable d’éprouver quelque sentiment que ce soit. Ces éléments hasardeux seront mis de côté dans cette recherche, car ils se fondent sur un jugement trop subjectif.
Bergeret (Encyclopédie médico-chirurgicale, 1980) tente de situer les caractères psychosomatiques souvent associés au comportement alexithymique : « un affaiblissement du fonctionnement de la pensée au profit d’un mode d’expression passant essentiellement par le corps. »
L’analyse des dysfonctionnements émotionnels sur le terrain clinique est relativement inexploitée. La pensée opératoire et l’alexithymie, sont une illustration exemplaire du rôle des conduites langagières et des représentations verbales dans l’élaboration d’une expérience émotionnelle. Il faut signaler l’importante différence entre un individu névrosé et un sujet alexithymique. Les premiers parviennent à refouler ou à se défendre contre des sentiments et des fantasmes associés au conflit psychologique, tandis que les seconds semblent avoir une lacune dans leurs possibilités d’avoir prise sur leurs sentiments intérieurs.
En bref, les critères cliniques de l’alexithymie sont ainsi présentés :
Anarthrie :
L'anarthrie est un trouble articulatoire rencontré notamment dans certains types d'aphasie. Elle est caractérisée par des difficultés articulatoires. Chez tous ces patients le cortex insulaire (l'insula) caché au fond de la scissure de Sylvius sous la région de Broca, est détruit.
Anhédonie : L'anhédonie est un symptôme médical retrouvé dans certaines pathologies psychiatriques et parfois chez le sujet exempt de trouble. Il caractérise l'incapacité d'un sujet à ressentir des émotions positives lors de situations de vie pourtant considérées antérieurement comme plaisantes. Cette incapacité est fréquemment associée à un sentiment de désintérêt diffus. L'anhédonie, perte de la capacité à ressentir des émotions positives, est fréquemment observée au cours de la dépression et de la schizophrénie.
Anosognosie (A Damasio "L'erreur de Descartes") :
l’anosognosie est un trouble neuropsychologique qui fait qu'un patient atteint d'une maladie ou d'un handicap ne semble pas avoir conscience de sa condition. A l'inverse du déni, qui est un mécanisme de défense psychologique "normal", cette méconnaissance par l’individu de sa maladie est pathologique et peut refléter une atteinte de certaines aires cérébrales. Elle est présente particulièrement dans certains types d'accidents vasculaires cérébraux, dans le syndrome de Korsakoff ou durant l'évolution de certaines maladies neurodégénératives. Elle peut notamment causer l'ignorance de certaines conditions graves comme une cécité ou une paralysie. Bien que ce trouble fut décrit dès 1914 par le neurobiologiste Joseph Babinski, sa cause reste encore mal comprise.
Le terme d'anosognosie est issu du grec nosos, "maladie" et gnosis, "connaissance". Le préfixe a- privatif donne à l'ensemble la signification d'absence de conscience de la maladie.
Les patients atteints d'anosognosie ont besoin d'être mis de façon flagrante en face de leur handicap pour qu'ils se rendent compte que quelque chose ne va pas chez eux.
Ces patients ne perçoivent donc pas le handicap qui les affecte. Le système sensoriel qui permettrait normalement cette perception, de façon automatique, rapide et interne, est donc, chez eux, dans l'impossibilité de fonctionner. Ce déficit est constant dans les cas sévères d'anosognosie, bien que dans les cas moins graves il puisse être masqué. Par exemple, un patient donné peut se souvenir avoir vu son bras inerte et, par déduction, admettre que quelque chose ne va pas dans cette partie de son corps. Ou bien tel autre patient peut se rappeler les innombrables déclarations, faites par ses proches ou les membres de l'équipe soignante, selon lesquelles il y a paralysie, il y a maladie, et que non, tout n'est pas normal. Se référant à cette information obtenue de source externe, l'un de nos plus intelligents patients atteint d'anosognosie, dit constamment : « J'ai eu autrefois ce problème », ou bien : « J'ai été autrefois affecté du syndrome de la dénégation. » Bien entendu, il continue à en être affecté. Cette incapacité à prendre conscience, dans l'immédiat, de l'état actuel du corps est absolument étonnante.
Outre la dénégation de leur maladie, les patients atteints d'anosognosie se caractérisent par un autre trait remarquable : ils ne se font aucun souci pour leur état ; ils n'expriment aucune émotion, et disent ne pas en ressentir, lorsqu'on les questionne à ce sujet. Lorsqu'on les informe qu'ils ont subi une grave attaque cérébrale, qu'ils sont menacés de façon imminente par une nouvelle hémorragie cérébrale ou bien qu'ils souffrent d'un cancer généralisé qui a maintenant gagné le cerveau – en bref, lorsqu'on leur apprend que leur vie ne sera plus jamais comme avant – ils reçoivent la nouvelle avec sérénité, faisant preuve quelquefois d'humour noir, mais ne montrent jamais angoisse ni tristesse, chagrin ni colère, désespoir ni panique. Il est important de se rendre compte que si vous communiquez ces mêmes mauvaises nouvelles à un patient atteint d'une lésion affectant l'hémispère gauche de façon symétrique, la réaction de ce dernier sera tout à fait normale. Les patients anosognosiques, quant à eux, sont totalement incapables d'exprimer et de ressentir de l'émotion, et c'est peut-être le seul aspect heureux de leur état par ailleurs tragique. Il n'est peut-être pas surprenant que, dans ces conditions, ils aient beaucoup de mal à faire des projets dans le futur, ainsi que de prendre des décisions, dans le domaine personnel ou dans celui des relations sociales. La paralysie est peut-être le moins grave de leurs problèmes.
Les patients atteints du type d'anosognosie décrit ci-dessus présentent une lésion dans l'hémisphère droit. Bien que la recherche des corrélats neuro-anatomiques de cette pathologie ne soit pas terminée, il est clair que l'atteinte en question touche certaines aires corticales particulières de l'hémisphère droit, appelées somatosensorielles (de la racine grecque acusga, pour « corps » ; le système somatosensoriel concerne aussi bien les sensations externes, comme celles relatives au toucher, à la température, à la douleur, que les sensations internes, comme celles relatives à la position des articulations, l'état viscéral et la douleur). Ces aires comprennent le cortex de l'insula ; les aires cytoarchitectoniques 3, 1 et 2 (dans la région pariétale) ; et l'aire S2 (également pariétale, dans la profondeur de la scissure sylvienne). (Remarquez que, lorsque j'emploie le terme de somatique ou de somatosensoriel, je vise tout le soma, c'est-à-dire le corps, dans le sens général, et que j'envisage tous les types de sensations corporelles, y compris viscérales.) La lésion frappant les patients anosognosiques affecte aussi la matière blanche de l'hémisphère droit, ce qui a pour effet d'interrompre les connexions reliant les différentes régions mentionnées ci-dessus (lesquelles reçoivent des messages en provenance de tout le corps –muscles, articulations, organes internes), ainsi que les
connexions les reliant au thalamus, aux ganglions de la base du télencéphale, et aux cortex moteurs et prémoteurs. Une lésion n'affectant que partiellement le système aux multiples composantes dont je viens de parler, ne provoque pas le type d'anosognosie que j'ai décrit.
J'avance depuis longtemps l'hypothèse que les aires cérébrales qui sont interreliées au sein de la vaste région de l'hémisphère droit lésée dans le cadre de l'anosognosie, engendrent probablement, par le biais de leur interaction, la carte la plus complète et la plus synthétique sur l'état du corps à chaque instant, dont puisse disposer le cerveau.
Une dominance se rapportant à l'hémisphère droit concerne la perception des informations sensorielles en provenance du corps : la représentation de l'état fonctionnel des viscères, d'une part, et celle de l'état fonctionnel des muscles squelettiques des membres, du tronc et du visage, d'autre part, se combinent en une carte dynamique coordonnée. Plus exactement, il n'y a pas combinaison en une seule carte, mais plutôt interaction et coordination entre signaux arrivant au niveau de plusieurs cartes distinctes. Dans ce cadre, les messages relatifs aux côtés gauche et droit du corps trouvent leur maximum de convergence dans l'hémisphère droit, au niveau des trois régions du cortex somatosensoriel mentionnées ci-dessus. De façon curieuse, la représentation de l'espace en dehors du corps, de même que les processus émotionnels, font l'objet d'une dominance hémisphérique droite. Cela ne veut pas dire que le corps ou l'espace n'est pas représenté dans les structures équivalentes de l'hémisphère gauche. Simplement, les représentations sont différentes : à gauche, elles sont probablement partielles, et ne font pas l'objet d'une intégration fonctionnelle.
Sous certains aspects, les patients atteints d'anosognosie ressemblent à ceux souffrant de lésions préfrontales. Par exemple, les patients anosognosiques sont incapables de prendre les décisions appropriées dans le domaine personnel ou dans celui des relations sociales, exactement comme les patients souffrant de lésions préfrontales. Et ces derniers, tous comme les anosognosiques, sont généralement indifférents à leur état de santé et semblent tolérer la douleur de façon inhabituelle.
Les anosognosiques du type que je décris ici ne souffrent donc pas seulement d'une paralysie du côté gauche, dont ils ne sont pas conscients. Ils présentent aussi un déficit dans les domaines du raisonnement et de la prise de décision, ainsi que dans celui des émotions et des sentiments.
Les observations sur les états pathologiques décrits ci-dessus, se traduisant de façon prépondérante par des déficits dans la faculté de raisonnement et de prise de décision, ainsi que dans la capacité d'exprimer et ressentir des émotions, mettent donc en évidence les faits suivants :
En bref, il semble bien y avoir dans le cerveau humain une série de systèmes neuraux impliqués de façon étroite dans les processus de pensée orientés vers un but, que nous appelons raisonnement, ainsi que dans l'organisation des réponses que nous appelons prises de décision, l'accent étant mis dans ces deux cas sur le domaine personnel et celui des relations sociales. Cette même série de systèmes est aussi impliquée dans les processus présidant à l'expression et à la perception des émotions, et se rapporte en partie au traitement des messages provenant du corps.
Conclusion :
Aphasie :
perte ou trouble du langage. Quatre modalités du langage peuvent être touchées : parler, comprendre, lire, écrire. 80% des aphasies sont dues à des attaques cérébrales : tromboses ou hémorragies. Autres causes : tumeur, traumatisme lors d'accidents de la route...
Aphasie de Broca:
Les patients parlent peu et mal mais comprennent bien.
Aphasie de Wernicke :
Les patients comprennent mal et parlent abondamment mais pas normalement.
Aphasie de conduction (de Lichtheim) :
Le patient comprend normalement mais jargonne comme un aphasique de Wernicke. En plus il montre de grandes difficultés à répéter les mots ou les phrases qu'on leur fait entendre, ce qui s'explique tout naturellement par la déconnexion entre compréhension et production de la parole.
Aujourd'hui on sait que le langage est le produit d'une mécanique cérébrale extrêmement compliquée et subtile et peut donc tomber en panne de mille manières différentes. Parler de l'aphasie de Broca de Wernicke ou de Licheim, entités aux coutours flous, comme entitées bien définies résultant d'une cause précise ne vaut guère mieux que regrouper sous une même étiquette toutes les pannes qui empêcheraient une navette spatiale de décoller ou de voler droit.
Aphasie transcorticale motrice :
Si l'on déconnecte le centre des concepts de celui de la mémoire motrice des mots, on peut prédire l'existence de patients dont la mécanique du langage fonctionne, qui peuvent répéter et comprendre, mais qui ne font pas usage de leur langage pour exprimer leurs idées.
Les régions cérébrales qui interviennent dans la production du discours sont encore mal connues et peu étudiées en neuro-imagerie. Par contre, la neuropsychologie fournit des exemples de lésions cérébrales qui semblent affecter spécifiquement cette capacité. L'exemple le mieux documenté est celui des lésions du cortex frontal latéral qui causent un trouble appelé aphasie transcorticale motrice (ATCM). Comme toutes les aphasies, l'ATCM est un trouble du langage ; mais les patients qui en souffrent sont capables de répéter des mots et même des phrases. Ils peuvent aussi nommer un objet qui leur est montré, lire un texte à haute voix et même fournir des réponses très courtes à des questions du type : « Où étiez-vous hier ? » Leur handicap concerne la génération du discours : ils n'arrivent pas à élaborer eux-mêmes des phrases. Leur trouble ne semble affecter aucune des composantes individuelles du langage, comme la sémantique, la phonologie, l'articulation, la grammaire ou la concaténation ni sa compréhension. L'ATCM est donc décrite comme un problème de planification de l'action dans le domaine du langage.
Sur le plan anatomique, ce trouble survient souvent après des lésions des parties les plus en avant du cortex frontal, en avant de l'aire de Broca, et parfois de la partie ventrale et latérale — en bas et sur le côté — du cortex préfrontal, qui fait partie du réseau par défaut. Cette position très antérieure n'est pas très surprenante compte tenu du parallélisme existant entre les systèmes neuronaux chargés de la production du langage et des comportements moteurs. Il existe de nombreuses similarités entre les mécanismes neuronaux permettant de bouger et de parler ; après tout, parler, c'est enchaîner des programmes moteurs articulatoires et donc des séries d'action, pour bouger la langue, la bouche et la gorge. Dans le cas du langage des signes, le lien entre langage et motricité est encore plus évident. Ces programmes articulatoires doivent obéir à des règles, grammaticales notamment, et s'adapter au contexte, pour atteindre un but, qui consiste à communiquer une information ou simplement produire un effet sur son interlocuteur. De la même façon, le comportement moteur, comme celui mis en jeu pour découper une tranche de pain, allumer la télé ou tirer un coup franc, est aussi un enchaînement d'actions, soumis à des règles, qui doit s'adapter au contexte pour atteindre un but. Ce but peut d'ailleurs être de véhiculer un message, ou une émotion, exactement comme le ferait la parole — songez au pianiste ou encore au danseur.
Aphasie transcorticale sensorielle :
Si on coupe la connexion entre le centre de la reconnaissance des mots et le centre des concepts, le patient peut répéter tout ce qu'il entend sans rien y comprendre.
Apraxie :
trouble des mouvements volontaires.
Apraxie idéatoire :
les patients chez qui la zone AIP (antérieure intrapariétale) située près du pouce du gant de boxe, est endommagée, souffrent souvent d'un trouble appelé apraxie idéatoire, ce qui signifie qu'ils ne savent plus comment utiliser certains objets de la vie courante :
devant une bougie et une boite d'allumettes, le patient va par exemple frotter la bougie contre la boite pour essayer de l'allumer.
En un sens les neurones d'AIP ne font donc rien d'autre en tant normal que de constater et mémoriser les actions le plus souvent associées à chaque objet usuel. Cette mémoire suffit néanmoins à informer le cortex prémoteur du mouvement qu'il doit préparer pour utiliser l'objet situé devant soi, dès lors que l'AIP s'active.
Démence sénile :
La démence chez les seniors est appelée démence sénile ou sénilité, et est perçue comme étant un aspect normal et, quelque part, inévitable chez les personnes âgées, plutôt qu'une maladie spécifique. En 1907, une maladie organique connue sous le nom de maladie d'Alzheimer a été décrite. Elle a été associée à des changements microscopiques cérébraux.
Durant les périodes 1913-1920, la schizophrénie est définie, et le terme de démence précoce a été suggéré pour qualifier le développement de la démence de type sénile dès le plus jeune âge.
Dyslexie phonologique :
Les patients sont incapables de lire des pseudo-mot comme "CHIGNADORLE" alors même qu'ils lisent correctement les vrais mots. Cette dyslexie est le résultat d'une destruction de la voie de lecture par application de règles, la seule possible pour lire les pseudo-mots. Les vrais mots, eux, sont lus par la voie lexicale.
Dyslexie de surface :
Ici pas de difficulté à lire les pseudo-mots. La voie de lecture régulière est donc intacte. Qu'en est-il des vrais mots ? Cela dépend. Ceux dont la prononciation peut être déduite de leur orthographe comme "CARAMEL" ne posent pas de problème, ce qui est normal si l'on admet l'intégrité de la lecture régulière. En revanche, les mots irréguliers donnent lieu à des erreurs de régularisation. Un tel lira "OIGNON" "ouagnon". Cela s'explique naturellement si l'on admet que la voie de lecture lexicale est hors d'usage.
Épilepsie
Héminégligence : Certaines lésions cérébrales affectent spécifiquement l'attention. Ainsi, les patients atteints au niveau de la partie arrière du lobe pariétal droit souffrent souvent d'un syndrome d'héminégligence, une tendance à négliger la partie gauche de leur champ visuel, voire à « oublier » son existence. Ces patients ne sont pas aveugles — ils peuvent voir certains objets situés sur leur gauche —, mais ne prêtent aucune attention à ce qui se trouve de ce côté. Un patient héminégligent peut oublier de se raser un côté du visage ou oublier de manger ce qui se trouve à gauche de son assiette. D'autres lésions du lobe pariétal peuvent empêcher la personne de voir deux objets à la fois, comme dans le syndrome de Balint. Ces troubles sont considérés par les neurologues comme des dysfonctionnements de l'attention.
Lésion du complexe moteur supplémentaire : Si le cortex moteur n'exécute pas toutes les actions que lui propose le lobe pariétal c'est grâce à un garde-fou situé dans le cortex frontal qui filtre les propositions d'actions jugées trop farfelues. Quand ce garde-fou fonctionne bien, la plupart des propositions du lobe pariétal passent inaperçues et nous n'en avons même pas conscience. Mais il suffit qu'une des composantes du garde-fou déraille pour que la partie arrière du cerveau prenne le pouvoir sur le corps. C'est ce qui arrive aux malheureux atteints de lésions du complexe moteur supplémentaire, ou CMS, un ensemble de régions corticales situées juste en avant du cortex moteur dans le lobe frontal. Ces patients souffrent de syndromes étranges, comme celui du membre étranger - Alien Limb Syndrome. L'un de leurs membres cesse d'obéir pour prendre son autonomie et mener sa vie propre, il devient étranger.
Leucoencéphalite :
Le terme Leucoencéphalite désigne toute affection cérébrale caractérisée anatomiquement par des lésions inflammatoires prédominant au niveau de la substance blanche (leuco=blanc) des hémisphères cérébraux, et dont l'origine est exogène (par opposition à la leucodystrophie, dans laquelle le processus est d'origine endogène).
Phénylcétonurie :
La phénylcétonurie est une maladie génétique rare et grave en relation avec un trouble du métabolisme de la phénylalanine (acide aminé d'origine alimentaire).
Elle affecte un nouveau-né sur 16 000 et est responsable d'une arriération mentale (oligophrénie phénylpyruvique) progressive en l'absence de traitement approprié. En France et dans de nombreux pays, on procède à la naissance à un dépistage systématique de la phénylcétonurie.
La phénylcétonurie a été découverte par le médecin norvégien Ivar Asbjørn Følling, en 1934, lorsqu'il a observé que l'hyperphénylalaninémie (HPA) était associée à un retard mental. En Norvège, cette maladie est appelée maladie de Følling, d'après celui qui l'a découverte, le Dr Følling, qui fut un des premiers médecins à étudier la maladie au moyen d'une analyse chimique minutieuse. Ayant analysé ainsi l'urine d'un frère et d'une sœur retardés mentaux, il demanda à un grand nombre de médecins près d'Oslo de tester l'urine d'autres patients retardés mentaux. Cela amena à découvrir la même substance que celle qu'il avait trouvée chez 8 autres patients. Cette substance ne put être analysée chimiquement que de façon beaucoup plus rudimentaire qu'on ne peut le faire aujourd'hui. Ses tests lui montrèrent que les réactions donnaient naissance à du benzaldéhyde et de l'acide benzoïque, ce qui l'a mené à la conclusion que ce composé contenait un anneau de benzène. Des essais supplémentaires montrèrent que le point de fusion était le même que celui de l'acide phénylpyruvique, ce qui indiquait que c'était la substance qui se trouvait dans l'urine.
C'est en 1953 que Horst Bickel propose un traitement par régime alimentaire. Robert Guthrie créé son test permettant le dépistage de masse chez le nouveau-né en 1963.
Polioencéphalite :
La polioencéphalite (polio=gris) est une maladie infectieuse due à un virus (neurotrope: spécifique du système nerveux) de la poliomyélite, de la diphtérie, de la scarlatine.
Prosopagnosie :
incapacité de reconnaître les visages. (prosopon=visage)
Simultagnosie (par JP Lachaux) : Troubles de la reconnaissance de plusieurs objets par un patient qui n'identifie bien que les détails. Il s'agit d'une variété d'agnosie qui est l'impossibilité de reconnaître des objets, alors que les fonctions sensorielles (vision, audition, toucher, etc...) sont normales. Autrement dit ce terme est l'impossibilité pour un patient de voir plusieurs choses à la fois. La simultagnosie se rencontre dans les démences et contribue à une dépendance rapide des patients.
La simultagnosie pose une question quasi philosophique... et y répond : qu'est-ce qu'un objet ? Si le médecin trace deux traits verticaux côte à côte sur une feuille, le patient est incapable de les comparer pour déterminer lequel des deux est le plus long. Mais si le médecin relie ces deux traits verticaux par un troisième trait horizontal pour former un U, le patient arrive à faire l'exercice. Il n'y a maintenant plus qu'un seul objet, que le patient parvient à percevoir dans sa globalité.
Le simple fait de connecter deux objets entre eux aide le patient à les voir simultanément, à condition que ce lien graphique donne vraiment l'impression que les deux objets n'en font plus qu'un.
La notion d'objet nous semble aller de soi.
Achromatopsie :
perte totale ou partielle de la perception des couleurs. Les personnes continuent à voir les formes, les mouvements et le relief mais elles percoivent le monde dans des teintes grises plus ou moins foncées.
Agnosie :
difficulté à identifier les objets. (agnosia=ignorance)
Akinetopsie : L'akinétopsie ou agnosie visuelle du mouvement est une incapacité de percevoir le mouvement. La personne atteinte d'akinétopsie voit avec des arrêts sur image qui peuvent durer plusieurs secondes. Cela présente de grandes difficultés dans la vie quotidienne, par exemple pour traverser la rue ou pour pratiquer un sport.
Zeki a étudié un autre paramètre que la couleur : le mouvement. Suivant la même logique, il a cherché si des régions cérébrales étaient plus activées par des images en mouvement que par les mêmes images immobiles. Comme dans le cas de la couleur, une région bien définie est apparue, nommée V5, située cette fois à la face latérale et non à la face inférieure du cortex visuel. Que se passe-t-il en cas de destruction de cette région vouée à la perception du mouvement ? Il est assez facile de s'imaginer ce qu'est un monde sans couleurs, mais sans mouvement ? La perte de la vision du mouvement a bien été décrite, sous le nom
d'akinetopsie. La chose est extrêmement rare.
Citons quelques lignes de la description que donne Zihl des difficultés rencontrées par une patiente, Madame L. M., dont la région V5 avait été endommagée dans les deux hémisphères : « Le problème visuel dont se plaignait la patiente était une perte de la vision du mouvement dans les trois dimensions. Par exemple, elle avait du mal à verser du thé ou du café dans une tasse, car le liquide semblait gelé, comme une cascade de glace. De plus, elle ne pouvait s'arrêter de verser au bon moment, puisqu'elle était incapable de voir le mouvement du liquide s'élevant dans la tasse ou le pot. En outre, la patiente se plaignait d'avoir du mal à suivre une conversation, car elle ne pouvait voir les mouvements du visage de son interlocuteur, en particulier les mouvements de ses lèvres. Elle se sentait très inquiète et mal à l'aise quand elle se trouvait dans une pièce où se déplaçaient plus de deux autres personnes, au point de quitter les lieux aussitôt, expliquant elle-même que "les gens apparaissent soudainement ici ou là, mais je ne les ai pas vus bouger". La patiente souffrait des mêmes difficultés, plus marquées encore, dans les rues pleines de monde, lieux qu'elle évitait donc autant que possible. Elle ne pouvait traverser la rue, incapable qu'elle était de juger de la vitesse des voitures, et cela bien qu'elle pût sans peine identifier les voitures elles-mêmes. "La première fois que j'aperçois la voiture, elle me paraît loin. Mais ensuite, au moment où je veux traverser, la voiture se trouve soudain toute proche." Peu à peu, elle apprit à estimer la distance des véhicules en mouvement en se fondant sur leur bruit, de plus en plus fort. »
Hémimicropsie :
Madame Hélène O., âgée de 60 ans, était secrétaire, retraitée depuis peu. Quelques jours après une opération bénigne de la cloison nasale, elle commença à se plaindre de troubles visuels étranges. Les objets et les visages semblaient disparaître de sa vue, mais pas entièrement puisqu'elle continuait à en voir des fragments. Par exemple, elle nous raconta qu'elle avait vu, et parfaitement reconnu, la boucle d'oreille que sa fille portait alors même qu'elle ne voyait pas le reste du visage de sa fille ; ou encore qu'elle avait vu un portemanteau fixé au mur face à son lit, juste entre deux fenêtres, sans pour autant voir ces deux fenêtres ; ou enfin qu'elle s'était levée de son lit, puis s'était retournée pour se recoucher, mais n'avait plus réussi à retrouver ce lit, qui avait comme disparu de la scène. Madame O. avait aussi remarqué qu'elle n'arrivait pas à attraper les objets qu'elle essayait de saisir : sa main passait presque toujours à côté du verre lorsqu'elle voulait boire. En quelques semaines, ces troubles, sur lesquels nous reviendrons, disparurent.
Elle s'aperçut alors d'une autre anomalie curieuse : lorsqu'elle regardait quelqu'un bien en face, l'oeil gauche lui semblait toujours un peu plus petit que l'oeil droit. Le même phénomène se produisait avec tous les objets symétriques : une paire de mains, un écouteur de téléphone, etc. La moitié droite paraissait toujours un peu plus petite que la gauche. Pour mesurer le plus objectivement possible ce phénomène, nous lui avons montré des paires de cercles, un du côté droit et un du côté gauche de son champ visuel, de tailles légèrement différentes, en lui demandant de nous montrer le plus grand des deux. Lorsque le cercle de droite était plus grand que celui de gauche de 5 %, Madame O. choisissait au hasard un cercle ou l'autre : son illusion visuelle, en compensant la différence de 5 % entre les deux cercles, les lui faisait paraître identiques. Nous avons observé le même phénomène (à ceci près que c'était cette fois la moitié gauche qui était rétrécie) chez un professeur de dessin, dont le talent artistique lui permettait de reproduire fidèlement ce qu'il percevait ; il agrandissait la moitié gauche des objets symétriques, comme les visages, afin qu'ils lui paraissent symétriques. Ces deux patients présentaient une petite lésion de la face latérale d'un lobe occipital, dans l'hémisphère opposé au rétrécissement visuel. Cette lésion épargnait les aires visuelles primaires, que nous connaissons déjà, et laissaient donc le champ visuel intact.
Métamorphopsie :
Trouble de la vision se caractérisant par une déformation des images.
Régions visuelles V4 et V5 : (Laurent Cohen)
Syndrome de Charles Bonnet :
Charles Bonnet naturaliste et philosophe Suisse est le premier à avoir décrit ces symptomes en 1760 dont souffrait son grand-père. Les sujets ont des hallucinations mais sont conscients qu'il s'agit d'allucinations et ce n'est que très récemment (1998) qu'on a pu mettre en évidence une hyperactivité du cortex visuel (cortex occipital et inféro-temporal) en rapport avec les hallucinations.
Ces hallucinations peuvent également provenir quand se forme un hématome dans cette région suite à la rupture d'un petit vaisseau cérébral.
NEUROTRANSMETTEURS
Plus de 50 neurotransmetteurs agissent dans le cerveau. De plus, leurs effets diffèrent selon la zone d'activation. Voici les plus importants :
Acétylcholine : C'est le premier neurotransmetteur qui a été découvert. Elle entre en jeu dans les aires du cerveau associées à la mémoire, l'attention, l'apprentissage ; elle est aussi impliquée dans la colère, l'agression, la sexualité et la soif. On note d'ailleurs une carence en acétylcholine chez les patients atteints de la maladie d'Alzheimer.
Elle permet également la jonction entre les motoneurones et les muscles.
Son absence provoque une nécrose de l'hippocampe et par voie de conséquence une amnésie de type Korsakoff, avant que les malades ne glissent lentement vers la démence, c'est la tristement célèbre maladie d'Alzheimer. L'acétylcholine a deux « serrures » possibles sur la membrane du neurone, des récepteurs muscariniques et des récepteurs nicotiniques. Vous avez bien lu, nicotinique comme nicotine ; la nicotine de la cigarette doit donc ses effets stimulants au fait qu'elle est une fausse clé pour les récepteurs de l' acétylcholine.
L'acétylcholine transmet des signaux depuis le nerf vague vers le coeur. L'ACh est également la substance qui transmet des messages depuis les nerfs périphériques vers les muscles, afin que ceux-ci se contractent. Le poison dans lequel certaines tribus indiennes enduisent leurs fléchettes contient une substance appelée curare, qui bloque les récepteurs ACh des fibres musculaires et paralyse ainsi la victime.
Il existe également des fibres cholinergiques qui vont du SNC vers la glande surrénale, qui sécrète entre autres l'hormone
appelée adrénaline. Cette hormone permet au corps de se battre - ou de fuir - en cas de situations de stress. D'autres fibres cholinergiques vont vers l'intestin, la vessie, etc. et ont un effet désactivant (parasympathique).
On sait aujourd'hui que deux sortes de récepteurs peuvent être stimulés par l'ACh. Certains récepteurs ACh réagissent à la muscarine (un composant de l'amanite, l'Amanita muscaria), d'autre pas. Ces derniers par contre réagissent à la nicotine, l'alcaloïde du tabac. Les récepteurs des fibres musculaires et de la glande surrénale sont du type nicotine, tandis que les récepteurs parasympathiques sont généralement du type muscarine. Tous les récepteurs ACh du SNC sont nicotinergiques. L'effet stimulant de la nicotine repose sur la stimulation de ces récepteurs.
Le pinang, un stimulant que l'on mâche dans de grandes parties du Sud-Est asiatique, contient de l'arécoline, qui agit sur les deux types de récepteurs.
Adénosine : L'adénosine - molécule indispensable à la synthèse de l'ATP, principale source d'énergie de notre organisme - est un neuromodulateur du système nerveux central qui possède des récepteurs spécifiques. Quand l'adénosine se fixe sur ses récepteurs, l'activité nerveuse est ralenti - nous devenons somnolents. L'adénosine facilite donc le sommeil et dilate aussi les vaisseaux sanguins, probablement pour assurer une bonne oxygénation lorsque nous dormons.
Effets de la caféine : les récepteurs à l'adénosine : animation
Adrénaline : Cette molécule porte aussi le nom d’épinéphrine. Elle agit comme un stimulant physique et mental. Elle accélère la vitesse de la respiration, dilate les pupilles et accroît le rythme cardiaque. Mais en excès, elle rend nerveux et peut entraîner la paranoïa.
Adrénaline et mémoire : Joseph Ledoux (Le cerveau des émotions)
Mais comment d'abord une situation émotionnelle peut-elle conduire à la libération d'adrénaline ? Cela, bien entendu, nous ramène à l'amygdale. Comme nous l'avons déjà vu maintes fois, cette région, quand elle détecte une situation d'aversion émotionnelle, met notamment en branle le système nerveux autonome. Il en résulte l'activation des surrénales qui libèrent alors l'adrénaline dans le sang. Celle-ci exerce son influence sur le cerveau, quoique de façon indirecte. Cette rétroaction, à la manière de William James, opère sur des systèmes également actifs à ce moment-là, comme l'hippocampe à l'origine du souvenir explicite de la situation. Bien que la manière dont cette rétroaction consolide la mémoire explicite ne soit pas encore complètement élucidée, il semble que l'adrénaline puisse en quelque sorte gagner le cerveau et influencer le fonctionnement du système de la mémoire du lobe temporal, renforçant les souvenirs qui y sont créés.
Les études de McGaugh et ses collaborateurs ont suggéré que l'adrénaline, une hormone libérée durant le stress, stabilise et renforce les souvenirs. Mais comme l'adrénaline ne peut normalement pas entrer dans le cerveau par la circulation sanguine (ses molécules sont trop grosses pour franchir la barrière hématoencéphalique), elle a une action indirecte. Le schéma montre comment elle pourrait agir indirectement sur le cerveau. Des stimuli associés à un danger activent l'amygdale. Le système nerveux autonome (SNA) est activé via l'hypothalamus latéral (HYP. LAT) puis la médulla latérale ventrale rostrale (LVR). L'un des nombreux organes à répondre à l'activation du SNA est la médullo-surrénale. Celle-ci libère l'adrénaline, qui a une large action dans l'organisme. Son effet sur le nerf vague est particulièrement important pour la modulation de la mémoire, car celui-ci se termine dans le noyau du tractus solitaire (NTS) du bulbe rachidien. Le NTS envoie ensuite des signaux au locus coeruleus (LC) qui libère de l'adrénaline dans de larges aires du cerveau antérieur, y compris l'amygdale et l'hippocampe. En influençant les fonctions de l'amygdale et de l'hippocampe, les souvenirs émotionnels implicites et les souvenirs explicites des émotions pourraient ainsi se trouver modulés.
Dopamine : Elle contrôle la stimulation de plusieurs zones du cerveau, et joue en particulier un rôle primordial dans la motivation physique.
Une importante carence en dopamine (comme dans la maladie de Parkinson), peut rendre impossible certains mouvements. Inversement, un excès de dopamine déclencherait des hallucinations et un état schizophrène. C'est notamment le mécanisme impliqué avec la cocaïne, qui empêche la recapture de la dopamine et accentue donc son action. La nicotine provoque aussi une augmentation de la transmission dopaminergique.
Animation sur Internet concernant la dopamine et le tabac
La dopamine n'est synthétisée que par quelques centaines de milliers de cellules, mais elles jouent un rôle extrêmement important dans les zones supérieures du SNC. On distingue parmi ces neurones dopaminergiques trois sous-groupes aux propriétés bien définies. Le premier groupe contrôle les mouvements : une carence en dopamine dans le système (nigro-striatal) se traduit par exemple par la maladie de Parkinson, dont les symptômes sont des tremblements, des raideurs et d'autres troubles moteurs, et, à un stade ultérieur, des signes de démence. Le deuxième groupe, le système mésolimbique, contrôle le comportement émotionnel. Le troisième groupe, le système mésocortical, projette exclusivement sur le cortex préfrontal. Dans cette partie de l'écorce cérébrale se situent différentes fonctions cognitives (mémoire, planification du comportement, abstraction), mais aussi plusieurs aspects émotionnels, en particulier liés au stress. Le système de gratification, que nous avons déjà abordé, fait partie de ce groupe. Le nucleus accumbens constitue un point de liaison important. La schizophrénie est liée aux troubles de ces deux derniers systèmes.
La dopamine semble intervenir davantage dans la motivation que dans la sensation de plaisir proprement dite. Si l'on abaisse artificiellement le niveau de dopamine chez des souris équipées du dispositif d'Olds et Milner, celles-ci continuent d'actionner spontanément la pédale, mais seulement si l'effort à fournir est minime. Le cerveau de ces souris reste donc sensible à la récompense déclenchée par l'action de la pédale, même sans dopamine. Des molécules connues pour les sensations de plaisir qu'elles procurent, comme la nicotine, l'éthanol et les cannabinoïdes présentes dans le tabac, l'alcool et Le cannabis, produisent cet effet même dans un cerveau dépourvu de dopamine. Un animal ou un être humain sans dopamine peut donc se réjouir d'une belle récompense, mais il ne fera aucun effort pour l'obtenir ; en un mot, il n'est pas motivé. Sans dopamine, je n'irai pas jusqu'au réfrigérateur pour me chercher une bière, même par 40 °C à l'ombre ; par contre, je serai ravi si vous allez m'en chercher une. C'est toute la différence entre ce que les chercheurs anglo-saxons appellent le wanting, le fait de vouloir quelque chose et d'être prêt à agir pour l'obtenir, et le liking, le fait d'apprécier cette même chose. S'il vous arrive parfois, le soir, d'être fatigué au point de ne pas parvenir à vous décoller du canapé pour alter au lit, tout en sachant pertinemment à quel point vous y seriez bien, vous avez une petite idée de ce que ressent un animal privé de dopamine.
Voir aussi le Circuit du plaisir et notamment le paragraphe : Un chemin vers le plaisir - Récompense et motivation (J-P Lachaux)
Endorphine ou Endomorphine : Découvertes dans les années 1970, les endorphines (de morphine et endogène) sont des neurotransmetteurs agissant sur les récepteurs opiacés. On les retrouve entre autres dans le cerveau et la moelle épinière ainsi que dans le système digestif.
Elle atténue la douleur, diminue la nervosité et donne une sensation de bien-être. Mais elles ralentissent aussi certaines fonctions vitales comme la respiration et entraîne des dépendances. C'est notamment la substance stimulée par les drogues comme l'opium ou l'héroïne. Mais on a noté que le sucre ou les graisses libèrent aussi de l'endomorphine.
Les premières endorphines ont été découvertes par John Hughes et Hans Kosterlitz dans le cerveau d'un cobaye. Ils appelèrent celles-ci enképhalines (du grec enkephalos: dans la tête).
Endorphine est un terme résultant d'une abréviation de l'expression substance morphinique endogène. En effet, les endorphines agissent comme la morphine, mais sont secrétées par l'organisme.
D'un point de vue purement scientifique être amoureux, c'est être stressé ce qui active notre amygdale ; hors elle ne se déclenche généralement que lorsqu'un danger nous menace en nous plaçant en état d'alerte (d'où l'agitation ressentit quand nous sommes amoureux). Dans le même temps l'état de panique nous rend aussi un peu aveugle ce qui explique pourquoi nous ne voyons pas vraiment les défauts de l'autre. Parallèlement les centres de la récompense (noyau acumbens) réagissent eux aussi et l'endorphine (un opiacé naturel) , sécrétée par l'hypothalamus nous rend complétement euphorique, tout comme la sérotonine produite par le noyau de Raphé et grâce à laquelle notre humeur est au beau fixe. Ce cocktail agit comme une drogue et pendant cette phase nous sommes comme en état de manque quand l'autre est absent : l'amour rend accroc et indulgent.
Enképhaline : Au début des années 1970, les chercheurs ont découvert dans le cerveau une substance jusqu'ici inconnue, l'enképhaline. Ils se sont aperçus qu'elle agissait comme la morphine pour apaiser et parfois supprimer la douleur. L'enképhaline inhibe les impulsions électriques qui véhiculent le message douloureux de la cellule nerveuse vers le cerveau.
GABA : En 1976, Braestrup et Squire, ainsi qu'une équipe indépendante de Hoffman La Roche, ont découvert que le cerveau contient des récepteurs qui réagissent spécifiquement aux benzodiazépines. En cherchant ensuite le transmetteur naturel, ils ont trouvé un transmetteur qui est lié aux récepteurs de façon encore beaucoup plus étroite que le valium. Ils ont baptisé cette substance 'acide gamma-amino-butyrique' (abréviation américaine : GABA). Il s'agit d'un des neurotransmetteurs les plus répandus du SNC; pratiquement tous les neurones ont des récepteurs GABA. GABA est un neurotransmetteur inhibitoire. Sa fonction est donc d'inhiber toute une série de systèmes activants. Il existe deux types de récepteurs GAGA : GABA-Ô et GABA-13. Seuls les récepteurs GABA-6 sont stimulés par l'alcool, les benzodiazépines et les barbituriques, ce qui se traduit par une sensibilité moindre à d'autres stimuli.
L'effet anxiolytique de l'alcool est généré par les récepteurs GABA.
Effets de l'alcool
Des neurones responsables du sommeil existent dans le cerveau humain, dans une petite région à la base du cerveau, l'« aire pré-optique ventro-latérale ». Actifs durant le sommeil, inactifs sinon, ces neurones libèrent un neurotransmetteur qui leur est propre, le neurotransmetteur « Gaba ». À l'inverse, on a identifié, dans d'autres régions du cerveau, des neurones qui sont actifs durant l'état de veille et inactifs durant le sommeil.
Glutamate : Le glutamate est un neurotransmetteur excitateur ubiquitaire dans le système nerveux central. Il agit sur les récepteurs N-méthyl-D-aspartate (NMDA) et non-NMDA amino-3-hydroxy-5-méthyl-4- isoxazole propionic acid (AMPA) et les récepteurs du kainate, qui sont tous des canaux ioniques. Il agit également sur les récepteurs métabotropes qui sont eux couplés au système de transduction intracellulaire lié aux protéines G. Le récepteur NMDA est probablement le plus étudié dans les pathologies psychiatriques, et nécessite parallèlement au glutamate un coagoniste obligatoire, la glycine.
Toute la difficulté dans l’utilisation des voies glutamatergiques dans les troubles psychiatriques tient à deux aspects essentiels de la transmission glutamatergique : son caractère ubiquitaire tout d’abord : à la différence de systèmes dont l’organisation anatomique est plus localisée, à l’exemple du système dopaminergique, la transmission glutamatergique n’est pas systématisée. Néanmoins la plus forte densité de récepteurs glutamatatergiques est retrouvée dans le cortex cingulaire antérieur, dans la surface médiale du cortex préfrontal et dans les hippocampes ;
l’excès de stimulation glutamatergique est toxique : si la transmission glutamatergique est excitatrice, à l’excès elle déclenche une entrée massive de calcium dans les neurones, entraînant la mort cellulaire par apoptose. Donc l’équilibre est difficile à trouver : une stimulation est excitatrice mais peut conduire à la mort cellulaire.
C'est le neurotransmetteur le plus courant (1/3 des transmissions synaptiques). Une carence en glutamate entraîne donc des difficultés d'apprentissage et de mémorisation à long terme.
Parmi les neurotransmetteurs les plus répandus du SNC, le GABA a un effet inhibitoire, alors que le glutamate a un effet excitant. Dans le domaine psychopharmacologique, le sous-type du récepteur N-méthyl-D-aspartate (récepteur NMDA) est le plus important. Son activation augmente la sensibilité aux stimuli des autres neurotransmetteurs. L'alcool et la PCP inhibent l'effet du glutamate et diminuent donc la sensibilité aux stimuli.
Histamine : un des principaux neuromédiateurs de l'éveil. Il a une influence sur l'appétit, la vision, la salive.
L'histamine est aussi un neurotransmetteur synthétisé et libéré par les neurones appelés « neurones histaminergiques ». Elle est stockée dans des vésicules (mais le transporteur responsable de cette localisation n'a pas encore été identifié). Elle est libérée suite à un stimulus électrique et va se lier à des récepteurs post- ou présynaptiques.
Mélanocortine : L'hormone mélanotrope, ou mélanotropine ou mélanocortine (nom le plus utilisé) ou MSH (pour melanocyte stimulating hormone), est une hormone sécrétée par les cellules mélanotropes de l'hypophyse (lobe antérieur), les kératinocytes et les mélanocytes. Elle est le produit de maturation d'une pro-hormone, la POMC, par des enzymes spécifiques appelées pro-hormone convertases.
Elle semble jouer un rôle important dans la dépression.
Mélatonine : La mélatonine, souvent dénommée hormone du sommeil, est surtout connue comme étant l’hormone centrale de régulation des rythmes chronobiologiques, et d’un certain point de vue, de pratiquement l’ensemble des sécrétions hormonales, chez l’humain, chez tous les mammifères et semble-t-il chez la plupart des espèces animales complexes. Cette neurohormone est synthétisée à partir d’un neurotransmetteur, la sérotonine, qui dérive elle-même du tryptophane, un acide aminé essentiel. Elle est sécrétée par la glande pinéale ou épiphyse (dans le cerveau) en réponse à l’absence de lumière.
La mélatonine semble avoir de multiples fonctions, autres qu’hormonales chez l’humain et les mammifères, en particulier comme antioxydant. Elle semble aussi jouer un rôle dans le système immunitaire.
Monoxyde d'azote : Il existe aussi des neurotransmetteurs de motricité spécifiques à une fonction donnée, qui ne sont pas dans le cerveau. Pour anecdote, citons le neurotransmetteur « monoxyde d'azote », au niveau du pénis, qui permet l'érection. Le médicament « Viagra » qui a permis de lutter contre les troubles de l'érection, agit sur ce neurotransmetteur.
Noradrénaline : La substance noradrénaline joue un rôle important dans les réactions que l'on a face à des situations de stress. Elle rend plus alerte (cette activation du cerveau depuis le tronc cérébral s'appelle la 'réaction d'alerte') et permet au corps de fuir ou de se battre, par exemple en stimulant le battement cardiaque, la circulation sanguine et la respiration de telle façon que les muscles reçoivent plus d'oxygène, et en vidant la vessie et l'intestin. C'est le réflexe classique de 'mouiller sa culotte'. Enfin, la norédraline donne une sensation de réussite, de victoire imminente, très importante quand il faut fuir ou se battre pour sauver sa peau.
Elle joue un rôle dans la réaction de l'organisme au stress. On pense qu'un taux élevé de noradrénaline, qui est probablement d'origine génétique, peut se traduire par une hypersensibilité. Les enfants timides ont un taux élevé de cortisol et de noradrénaline dans leur sang. Un taux trop bas se traduit par des problèmes de concentration et par une
incapacité de 'faire la part des choses'. La noradrénaline a la fonction de neurotransmetteur dans le SNC, mais également d'hormone lorsqu'elle est sécrétée avec de l'adrénaline par la glande surrénale. La plupart des corps cellulaires des neurones noradrénergiques se trouvent dans le locus coeruleus, un noyau du tronc cérébral. Ces neurones envoient leurs axones vers le système limbique (inhibition de l'appétit), les noyaux subcorticaux et l'écorce cérébrale (l'état d'alerte). Le produit clonidine qui est utilisé contre les tensions artérielles trop élevées inhibe l'activité du locus coeruleus. Le même effet se retrouve dans les opiacés et les endorphines. Pendant la phase de sevrage, le locus coeruleus est hyperactif (l'inhibition de l'opiacé disparaît). Pour faciliter la désintoxication, on prescrit parfois de la clonidine, qui se charge de diminuer l'hyperactivité du noyau.
Les autres corps cellulaires noradrénergiques se trouvent en divers endroits du tronc cérébral et envoient une partie de leurs axones vers les amygdales et une autre vers les neurones qui règlent la tension artérielle.
Ocytocine : Considérons les « campagnols » sortes de rats des champs ; chez les campagnols de l'Illinois, le neurotransmetteur ocytocine est impliqué dans les contractions de l'accouchement et la lactation. C'est-à-dire que l'arrivée à maturation du foetus par l'organisme déclenche (probablement sous forme de synthèse de protéines) une sécrétion d'ocytocine, qui engendre elle-même des commandes, telles que les contractions et la production de lait.
Comment aimer des enfants qui ne sont pas les siens, ou au contraire, délaisser ses propres enfants.
Prostaglandine : Autre découverte chimique importante au XXe siècle, certains tissus du corps produisent des prostaglandines, qui aident l'organisme à transmettre les signaux de douleur, et donc vous aident à ressentir la douleur. Le pharmacologue britannique John Robert Vane (né en 1927) a démontré en 1971 que l'efficacité de l'aspirine est liée à sa capacité à inhiber la production de certaines prostaglandines.
Sérotonine : "Molécule du bonheur", elle a un effet essentiel sur l'humeur et l'anxiété : à concentration élevée, elle rend optimiste et serein. De nombreux anti-dépresseurs comme le célèbre Prozac renforcent donc son action. Elle aurait également des effets sur le sommeil, la douleur, l'appétit et la pression artérielle. L'Ecstasy et le LSD par exemple accroîssent fortement le taux de sérotonine.
Animation des ISRS (inhibiteurs sélectifs de la recapture de la sérotonine)
Les corps cellulaires des neurones qui travaillent avec de la sérotonine (également appelée 5-hydroxytryptamine, 5-HT) - les neurones sérotonergiques - se trouvent dans les 'noyaux du raphé' du tronc cérébral et envoient leurs prolongements (les axones) vers différentes zones de notre cerveau. Un des aspects les plus étonnants de ces neurones est qu'ils génèrent eux-mêmes des potentiels d' action, contrairement à la plupart des neurones. On peut les comparer à un pace maker, le dispositif qui génère des impulsions électriques pour aider le coeur à se contracter. Cette activité spontanée est ensuite modulée (accélérée ou ralentie) par un grand nombre d'autres neurotransmetteurs, dont la sérotonine. Il existe donc un système autorégulateur : la sérotonine est libérée au niveau des synapses mais inhibe ensuite la sécrétion de sérotonine. C'est ce que l'on appelle une rétroaction négative.
Les neurones sérotonergiques sont surtout liés à l'écorce cérébrale, au système limbique et à une série d'autres centres cérébraux qui ont une fonction régulatrice au niveau sensoriel, moteur et associatif. On entend par 'associatif' le fait que les influx provenant des différents systèmes reçoivent une cohésion interne. Un bel exemple d'association est encore une fois la 'langue', un phénomène qui associe entre eux les aspects auditifs (entendre), visuels (voir des objets et lire en voyant des lettres) et moteurs (le contrôle des muscles du larynx). Pour bien intégrer toutes ces fonctions, notre cerveau dispose d'une série de points de liaison appelés 'relais'. La sérotonine a un effet inhibitoire sur les relais sensoriels et excitant sur les relais moteurs. Remarquons enfin que les effets de la sérotonine sont relativement lents à se manifester et à disparaître, ce qui suggère qu'elle a surtout un effet modulateur sur une activité synaptique trop rapide.
Il semble que le système sérotoninique ait un puissant effet homéostatique sur la coordination de types d'activités sensoriels et moteurs complexes dans le cadre de types de comportements très diversifiés. Plus l'individu est alerte, plus le système sérotoninique est actif. Ou mieux : plus le système sérotoninique est actif, plus l'individu est alerte. Le système sérotoninique n'est inactif que dans la phase de sommeil REM, pendant laquelle le cerveau est parfaitement 'éveillé' et l'individu rêve le plus. Aussi étrange que cela puisse paraître, le système sérotoninique correspond bien à la situation de la phase REM : intense activité cérébrale et motricité fortement inhibée. Les syndromes psychiatriques sont de plus en plus souvent associés à des anomalies au niveau du système sérotinique : troubles affectifs, schizophrénie et états hyperagressifs. Il est important de constater dans ce cadre que l'effet de beaucoup d'antidépressifs semble être basé sur une intensification de l'activité sérotonergique. Les spécialistes pensent qu'il existe également un rapport entre une fonction sérotoninique inhibée et les troubles caractériels antisociaux, les comportements violents et les comportements impulsifs. La fenfluramine, un inhibiteur de l'appétit très fréquemment utilisé (nom de marque Ponderal), a un puissant effet négatif sur le système sérotoninique : une administration intraveineuse de 40 mg/kg sur des rats fait baisser le taux de sérotonine pendant deux semaines.
La sérotonine a été identifiée dans la muqueuse gastro-intestinale, dans les plaquettes sanguines et dans le système nerveux central. La sérotonine présente dans la muqueuse gastro-intestinale représente environ 80 % de la sérotonine totale de l’organisme.
L’activité des neurones du raphé est liée au cycle veille-sommeil. Au cours du sommeil, les neurones du raphé sont silencieux et ne libèrent donc pas de sérotonine. Lorsque le sujet est éveillé, les neurones du raphé produisent des potentiels d’action à intervalles très réguliers. La fréquence des influx nerveux est corrélée à l’activité motrice, ce qui suggère que la quantité de sérotonine libérée dans le système nerveux central est augmentée avec l’activité motrice.
Comme la sérotonine joue un rôle important dans les changements d’état émotionnel, on pense que certaines molécules analogues à la sérotonine peuvent modifier ces états émotionnels. Les inhibiteurs sélectifs de la recapture de la sérotonine (des substances qui bloquent la recapture de la sérotonine, par exemple la fluoxétine), sont utilisés en médecine pour soigner les symptômes liés à la dépression ou aux troubles obsessionnels compulsifs.
Mécanisme de conversion en sérotonine
Des études ont montré que la sérotonine joue un rôle primordial dans la dépression, l’humeur, l’anxiété, le sommeil, le contrôle de l’appétit, la mémoire, l’apprentissage, la régulation de la température, le comportement sexuel, les hallucinations, les fonctions cardio-vasculaires, les contractions des muscles squelettiques, la régulation des hormones endocriniennes, la coagulation du sang et la motilité du système gastro-intestinal.
C’est dans les années 1970 que l’on a pris conscience du rôle important de la sérotonine pour la dépression. Chez de nombreuses personnes souffrant de dépression, on avait observé de faibles niveaux d’un métabolite de la sérotonine, l’acide 5-hydroxyindoleactique (5-HIAA), dans le liquide entourant le cerveau et la moelle épinière. Cela pouvait signifier que le cerveau ne produisait pas, ne métabolisait pas des quantités normales de sérotonine3.
Des chercheurs ont également constaté que les personnes qui tentaient de se suicider avaient des niveaux anormalement bas de 5-HIAA. Ces données suggéraient qu’une déficience en sérotonine pourrait en fait prédisposer certaines personnes à se tuer.
On a également montré que la sérotonine joue un rôle important dans les comportements agressifs.
La sérotonine est ce neurotransmetteur que l'on associe fréquemment aux troubles de l'humeur. Le cerveau produit la sérotonine à partir du triptophane, présent entre autres dans le chocolat, dont on nous vante parfois les vertus antidépressives. En privant le cerveau de triptophane, le niveau de sérotonine présent dans l'amygdale baisse, et les neurones qui s'y trouvent deviennent plus réactifs aux stimuli négatifs, ceux jugés de mauvais augure. Ces stimuli ont alors tendance à devenir saillants : l'attention se laisse plus facilement capturer par ce qui est menaçant, ou triste, et le cerveau y est plus perméable. Cet effet disparaît quand le niveau de sérotonine présent dans l'amygdale augmente à nouveau. D'ailleurs, de nombreux médicaments contre la dépression agissent directement sur cette sérotonine. Normalement, la sérotonine qui vient de servir à transmettre l'information d'un neurone à un autre ne reste pas dans l'espace synaptique qui les sépare : elle est recapturée par le neurone qui l'a envoyée pour resservir la prochaine fois, grâce à un système de petites pompes. Ces médicaments ralentissent l'action de ces pompes et augmentent ainsi mécaniquement la quantité de sérotonine présente dans la synapse ; c'est le principe des inhibiteurs sélectifs de la recapture de la sérotonine, comme le fameux Prozac.
Animation des ISRS (inhibiteurs sélectifs de la recapture de la sérotonine)
Vasopressine : Suivant les espèces, les campagnols mâles peuvent être à tendance marquée polygame ou monogame. Or, on observe qu'une augmentation d'un neurotransmetteur, la vasopressine, dans une aire précise du cerveau (le pallidum ventral) d'un campagnol mâle « polygame » le rend monogame ! On constate également que, outre sa fidélité, la vasopressine augmente l'instinct paternel du campagnol mâle... Si maintenant, on diminue ce neurotransmetteur dans une autre aire du cerveau (l'amygdale médiane), on supprime le comportement d'attachement paternel aux petits : il ne les caresse plus, ne les lèche plus, etc.
ONYCHOPHAGIE
L'onychophagie est traitée seulement avec notre propre volonté, c'est-à-dire, sans aucune aide externe. Il n'existe pas seulement un traitement concret pour arrêter de manger ou se ronger les ongles. Le seul fait que vous êtes arrivé á cette page, représente presque 90% de l'effort que vous venez d’effectuer pour abandonner cette manie. Les 10% restants, il s'agit de dominer les nerfs, l'anxiété, ou les problèmes quotidiens qui vous entraînent à cette mauvaise habitude.
Avant tout vous devez :
Du fait que la prise de conscience est en rapport étroit avec la volonté, nous pouvons contrôler adéquatement nos actes ou actions comme nous voulons. En résumé, prendre conscience c’est nous rendre compte de tout ce qu'il nous arrive ou ce que nous faisons, et à la fois, avoir la volonté de modifier ou renoncer à nos conduites. Dans la mesure où nous prenons conscience chaque fois que nous portons nos doigts à la bouche, nous progresserons vers le dépassement de cette mauvaise habitude.
Faisons un essai : Portez votre doigt vers votre bouche et retenez le mouvement quand votre doigt arrivera à 5cm de l'objectif. Répétez cette action plusieurs fois et à des distances différentes. Maintenant essayez de mettre le doigt entre les dents une ou 2 minutes et sans envie de mordre votre ongle. Ce petit exercice qui vient d'être effectué se nomme autodiscipline. Tous ces actes vous avez pu les contrôler pendant quelques minutes consciemment et avec votre autodiscipline, et peu à peu vous verrez comment vous pourrez contrôler cette habitude et vous quitter le désir de ronger vos ongles POUR TOUJOURS !
Pour vous aider á laisser progressivement l'habitude de ronger les ongles, essayez aussi de mordiller une racine de réglisse ou d'autres plantes comestibles. Vos dents et votre bouche seront assez occupées avec ces petites ramilles pour ne pas accepter à d'autres hôtes comme sont vos doigts ou vos ongles. La racine de réglisse NE SE MANGE PAS elle se mastique lentement. L’important c'est de la garder entre les dents le plus longtemps possible et seulement dans les situations qui activent, provoquent ou stimulent votre habitude de mordre ou ronger les ongles.
Nous mentionnons la racine de réglisse car c'est une plante naturelle très connue, surtout par ses propriétés médicinales, les enfants aiment la mastiquer pour sa saveur douce. Nous pouvons aussi essayer de mastiquer d'autres plantes comestibles de chaque région ou pays, du moment que nous avons la bouche et les dents bien occupées un certain temps, jusqu'à pouvoir nous déshabituer peu à peu de cette mauvaise manie.
La racine de réglisse nous pouvons la trouver sur tous les marchés et herboristeries du monde. On ne doit jamais mastiquer plus d’une semaine cette plante ou autres plantes comestibles avant de vous renseigner en vous assurant que vous n’y êtes pas allergique, et qu’il n’y a pour vous aucune contre-indication á ces plantes médicinales. Lire information sur la racine de réglisse.
Et que cela reste clair : Les véritables coupables ne sont pas vos ongles, ce sont vos dents.
Autre procédé pour avoir les mains occupées : Il consiste à effectuer un exercice simple qui sera pratiqué dans tout lieu, dans le bureau, à la maison, etc. chaque fois que vous avez des envies de mordiller les ongles. Tambourinez doucement avec les bouts des doigts sur une surface quelconque. Ce mécanisme, en plus de vous aider à vous défaire du désir de ronger les ongles, les fortifie et il leur donnera résistance.
Il est aussi très important de se détendre physiquement et mentalement avec des pratiques d'exercices de relaxation et respiration, yoga, gymnastique, sports et s’entretenir avec un certain hobby, sortir souvent de la maison, voir des choses nouvelles et amis, etc. de cette manière vous aurez la possibilité de vous détacher de la monotonie et refouler vers l’extérieur votre agressivité ou frustration enfermées en vous.
SAILLANCE
par Jean-Philippe Lachaux : L'attention visuelle a un rythme, et également une vitesse : le phénomène de l'inhibition du retour est la preuve que l'attention ne se déplace pas à une vitesse infinie ; si elle est prise à contre-pied, c'est qu'elle a une certaine inertie. Trois chercheurs de l'Université d'Utrecht aux Pays-Bas ont calculé récemment que l'attention met 150 millisecondes - un sixième de seconde - pour se déplacer de 7 degrés d'angle
visuel, c'est-à-dire la distance séparant l'index de l'auriculaire quand vous étendez le bras devant vous avec Les doigts écartés, comme pour dire « stop ». Selon l'estimation de cette équipe, il faut donc un sixième de seconde pour que votre attention passe de l'index au petit doigt. Quand vous faites face à la rue, l'attention visuelle est donc plus lente qu'un vélo, ou même un joggeur qui passerait à 6 mètres de vous à plus de 15 km/h.
L'attention visuelle aime bouger, mais elle ne bouge pas très bien ni très vite. C'est une raison supplémentaire pour avoir équipé le cerveau d'un système d'analyse pré-attentive de l'environnement capable de guider l'attention vers les éléments a priori les plus intéressants. Ce système produit à tout moment une sorte de carte touristique indiquant la position des principaux points d'intérêt, un peu comme les cartes Michelin. Cette carte définit ce que l'on appelle la saillance, la capacité à attirer l'attention, c'est donc une carte de saillance, dont plusieurs éléments laissent penser qu'elle se situe, au moins en partie, dans le lobe pariéta110. Dans une carte de saillance neuronale, en effet, les neurones doivent s'activer en fonction de l'importance de ce qui leur est présenté, plus qu'en fonction de sa forme, de sa couleur ou de sa vitesse. C'est ce que l'on observe dans une région du lobe pariétal appelée LIP, correspondant à la partie latérale (L) du sillon intrapariétal (IP), d'où son nom. LIP fait partie du système visuel et les neurones y ont même un champ récepteur, mais leur activité ne signale pas simplement la présence ou l'absence d'une forme ou d'un mouvement particulier ; elle signale surtout, lorsqu'elle est élevée, que l'individu va orienter son attention vers cette région de l'espace. Autrement dit, il suffirait de mesurer dans votre cerveau l'activité de tous les neurones de LIP pour deviner, sans trop se tromper, où va s'orienter votre attention visuelle. Et comme la nature est bien faite, ces neurones sont voisins d'autres neurones de LIP très impliqués dans le déplacement du regard, ce qui permet au cerveau de coordonner facilement les déplacements du regard avec ceux de l'attention, avec l'aide d'une région sous-corticale répondant au joli nom de colliculus supérieur ; d'où, sans doute, le phénomène de capture oculomotrice.
Calculer la saillance
L'attention est totalement capturée par un stimulus lorsque le lobe frontal décide de s'impliquer pour l'analyser en détail et le mémoriser. Être saillant, c'est donc être capable de déclencher cette réaction en chaîne, ce qui passe la plupart du temps par une réponse forte et synchrone au sein des régions sensorielles – voir le chapitre précédent. La saillance d'un événement sensoriel dépend donc directement de l'ampleur et du degré de coordination de la vague d'activité neuronale qu'il suscite. Le stimulus qui génère la réponse neuronale la plus forte ou la mieux organisée attire à lui toute l'attention. C'est un mécanisme de type winner-take-all, ou « le gagnant remporte toute la mise ». Cette règle simple suffit à expliquer pourquoi un flash lumineux ou un coup de canon attire naturellement l'attention, puisque dans les cortex sensoriels, l'intensité de la réponse neuronale augmente directement avec le contraste et l'intensité du stimulus.
Si l'on s'en tenait aux mécanismes décrits jusqu'ici, la carte de saillance serait statique, et l'attention resterait irrémédiablement fixée sur le clignotant de la voiture garée en double file. C'est bien sûr faux : le clignotant est effectivement un stimulus très saillant, mais il perd vite son pouvoir de capture ; l'attention s'en détourne rapidement pour aller voir ailleurs. La carte de saillance évolue donc promptement pour rediriger l'attention vers des éléments encore inexplorés. Le phénomène d'inhibition du retour, que nous avons déjà évoqué, participe à cette évolution, en empêchant l'attention de revenir immédiatement sur une zone déjà explorée. À plus longue échéance, un autre phénomène appelé habituation contribue également à cette recherche de nouveauté : lorsqu'un neurone est confronté plusieurs fois à la même stimulation, sa réponse décroît progressivement. C'est le cas par exemple, si vous êtes assis au café en train de discuter avec un ami, et que votre attention s'arrête sur une grande photo de Marylin Monroe collée au mur derrière lui. À ce moment précis, l'image de Marylin génère une activité neuronale très forte dans votre voie visuelle ventrale jusque dans la pointe antérieure du lobe temporal, dans ce que l'on appelle le cortex entorhinal et périrhinal et dans une structure voisine joliment nommée hippocampe. Mais cette réaction va perdre en vigueur à chaque fois que votre regard se posera sur l'image, jusqu'à atteindre un niveau plancher insuffisant pour déclencher une réponse du lobe frontal : le phénomène d'habituation a rendu Marylin moins saillante.
D'une façon générale, le cerveau réagit plus vivement à ce qui est nouveau, et c'est ce qui explique en partie pourquoi l'attention aime tant la nouveauté. Naturellement, vous auriez eu beaucoup plus de mal à vous lasser d'un écran de télé, dont l'image change tout le temps.
Même en prenant en compte l'habituation et l'inhibition du retour, aucun programme informatique ne peut prédire avec exactitude sur quelle partie d'une image va se porter l'attention d'une personne. Les déplacements spontanés de l'attention dépendent de nombreux autres facteurs, dont certains varient d'un autre. Au-delà de la nouveauté, l'attention est plus facilement distraite par exemple par ce qui revêt un caractère émotionnel fort, en fonction, bien sûr, de la sensibilité de la personne. Ainsi, un visage en colère capture généralement plus facilement l'attention qu'un visage à l'expression neutre. C'est heureux, car les stimuli émotionnels se doivent d'attirer l'attention : nous ne pouvons pas nous permettre d'ignorer un serpent ou une personne furieuse s'avançant vers nous. Il est donc normal que la saillance d'un événement ou d'un stimulus dépende aussi de sa valence émotionnelle, positive ou négative. Et c'est bien le cas : après avoir soumis des dizaines de sujets à des photos de serpents, des aboiements de chiens agressifs, des visages ou des voix en colère et toutes sortes d'horreurs, les chercheurs sont arrivés à la conclusion que les sons et les images à forte valence émotionnelle génèrent au niveau des cortex sensoriels des réponses plus fortes que les stimuli neutres, même dans des conditions de laboratoire confortables où la personne sait qu'elle ne risque rien. La Fusiform Face Area, cette région du gyrus fusiforme spécialisée dans l'analyse des visages, réagit plus fortement à un visage quand il exprime la peur, même s'il ne s'agit que d'une petite photo sur un ordinateur'''.
Cette amplification de la réponse neuronale pour les stimuli émotionnels intervient très tôt, dans les deux premiers dixièmes de seconde suivant l'apparition du visage à l'écran, dans le cas du gyrus fusiforme. Cet effet n'est donc pas dû à une réorientation top-down de l'attention, de haut en bas, impulsée par le lobe frontal, puisque ce dernier ne réagit qu'au bout de deux ou trois dixièmes de seconde. Si cette réorientation a lieu, l'amplification liée au caractère émotionnel du stimulus en est donc la cause et non la conséquence ; elle agit pour en augmenter la saillance. Le cerveau sait donc reconnaître la valence émotionnelle d'une image ou d'un son avant même d'y faire attention. Et effectivement, les stimuli à fort caractère émotionnel sont plus saillants que les autres : il faut moins de temps pour trouver un visage perdu au milieu d'autres formes quand celui-ci exprime une émotion forte. Il faut aussi moins de temps pour repérer une araignée ou un serpent qu'une fleur ou un chat.
SYNDROME POST-TRAUMATIQUE
Il survient à la suite d'un stress extrême (guerre, catastrophe naturelle, autres)
- lire la suite
Le MFB, par le cycle « désir – action – satisfaction », et le PVS, par la réponse de fuite ou de lutte réussie, amènent tous les deux l’organisme à préserver son homéostasie par l’action et forment ce que l’on appelle le système activateur de l’action (SAA).
Il en va tout autrement d'un troisième circuit, le système inhibiteur de l’action (SIA) (ou « Behavioral Inhibitory System (BIS) » en anglais). La mise en évidence de ce système revient à Henri Laborit au début des années 1970. Il est associé au système septo-hippocampal, à l’amygdale et aux noyaux de la base. Il reçoit des input du cortex préfrontal et envoie ses outputs à travers les fibres noradrénergiques du locus coeruleus et par les fibres sérotoninergiques du raphé médian. Certains reconnaissent d’ailleurs un rôle majeur à la sérotonine dans ce système.
Le SIA est activé lorsque la lutte et la fuite apparaissent impossibles et que le choix d’un comportement ne se résume plus qu’à subir passivement. Les conséquences pathologiques de cette inhibition de l'action ont permis de comprendre à quel point un stress chronique peut devenir destructeur pour l’être humain.
Le SIA est le fruit d’une évolution où il a été utile en fonctionnant sporadiquement, empêchant temporairement toute action inutile qui ne pourrait qu’empirer la situation. Pensons par exemple au petit mammifère qui se retrouve en plein milieu d’un champ et aperçoit un rapace au-dessus de lui; la meilleure chose à faire est encore de ne pas bouger et d’espérer passer ainsi inaperçu.
Or dans nos sociétés basées sur la compétitivité, nombreuses sont les personnes qui activent de façon chronique ce circuit pour éviter des représailles. L’inhibition de l’action n’est plus alors qu’une simple parenthèse adaptative entre des actions d’approche ou de retrait, mais une véritable source d'angoisse. C’est ce mal-être qui va peu à peu miner la santé de l’individu.
En effet, les conséquences négatives de l’inhibition de l’action sont nombreuses et ont été abondamment décrites : dépression, maladies psychosomatiques, ulcères d’estomac, hypertension artérielle sont les plus évidentes. Mais des dérèglements génétiques plus graves comme les cancers et l’ensemble des pathologies associées à une diminution de l’efficacité du système immunitaire sont aussi susceptible de découler de l’activation prolongée du SIA.
En effet, cet état amène des remaniements hormonaux à l'origine entre autre de hauts taux de glucocorticoïdes sanguins dont l'effet néfaste sur le système immunitaire est bien connu. Voilà pourquoi demeurer trop longtemps dans un état d'inhibition de l'action peut ouvrir la porte à la dépression et même à toutes les pathologies par l'entremise de cet affaiblissement du système immunitaire.
Inversement, et c'est particulièrement le cas dans nos sociétés, un trop grand nombre d'informations que l'on n'a pas appris à classer et à situer dans les niveaux d'organisation qui les englobent (et qu'elles englobent) provoque aussi le sentiment de l'inefficacité de l'action. Ce sont les bribes d'information dissociées de leur contexte du téléjournal ou le bombardement incessant de la publicité.
Enfin l'imaginaire propre à l'être humain, en permettant d'élaborer des scénarios que l'individu redoute de voir se réaliser, peut lui aussi amener l'individu à être inhibé dans son action. Le rêve d'un amour idéal inassouvi en a ainsi rendu plus d'un mal dans sa peau…
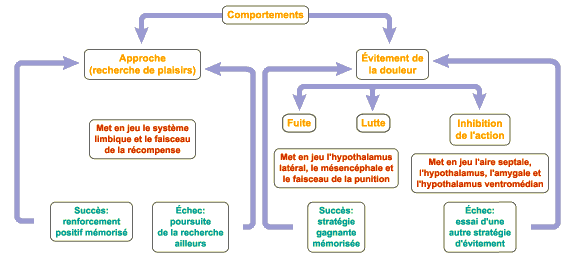
Au sein du circuit de la récompense, les neurones communiquent principalement grâce à deux types de neurotransmetteurs : celui des opioïdes et celui de la dopamine. Les opioïdes — qui doivent leur nom aux effets qu'ils produisent, semblables à ceux de l'opium — semblent liés à la sensation de plaisir et de déplaisir : le contact avec un stimulus plaisant ou déplaisant, quelle que soit sa modalité sensorielle, entraîne une libération d'opioïdes, dits opioïdes-mu. Et si l'on injecte ces opioïdes-mu directement dans le pallidum ou dans l'amygdale d'un animal recevant de la nourriture, celui-ci semble l'apprécier davantage. Inversement, le fait de bloquer l'action de ces opioïdes, avec une molécule appelée naloxone, diminue le plaisir ressenti : chez l'être humain, par exemple, La naloxone réduit considérablement la qualité de l'orgasme...
L'attention éclaire le monde et nos pensées, nos sensations et nos sentiments comme une torche. "Mon expérience est définie par ce à quoi je porte attention" disait William James.
La prochaine fois que vous pénétrerez dans un endroit familier, qu'il s'agisse de l'habitacle de votre voiture, de votre bureau ou de l'entrée du métro, observez les déplacements spontanés de votre attention. Vous remarquerez que cette dernière a tendance à viser toujours les mêmes endroits, selon un chemin stéréotypé. Il ne s'agit pas d'une loi absolue, mais d'une tendance. Nous avons tous des habitudes attentionnelles. Une personne à qui on demande de trouver un feu de circulation sur la photo d'un carrefour commence spontanément par chercher dans la partie droite de l'image. C'est une stratégie normale et efficace, puisque c'est là que se situent en général les feux. C'est aussi la preuve que les déplacements de l'attention tiennent compte de connaissances a priori concernant l'organisation spatiale de notre environnement. La plupart des environnements dans lesquels nous évoluons ont une organisation spatiale stable qui nous est familière – chaque place a sa chose, et chaque chose a sa place –, si bien qu'avec l'expérience, nous finissons par apprendre où se trouvent les éléments importants. En arrivant en voiture à un carrefour, nous devons rapidement savoir si ce carrefour a un feu et si ce feu est vert. C'est donc vers lui que l'attention se portera en priorité, parce que notre cerveau en a pris l'habitude, même si le feu de circulation n'est pas nécessairement l'élément le plus saillant de la scène en termes de luminosité, de couleur ou de taille. De même, face à un portrait, le regard et l'attention se déplacent selon une trajectoire bien connue privilégiant les yeux et la bouche, les parties du visage qui permettent le mieux de reconnaître une expression de joie, de colère ou de tristesse.
Et ce n'est pas tout, car le cerveau dirige aussi l'attention en fonction du regard des autres : si la personne assise en face de vous dans le bus regarde derrière vous d'un air insistant, vous finirez sans doute par jeter un coup d'oeil. C'est ce que l'on appelle l'attention conjointe ou l'attention sociale, une forme d'orientation de l'attention par le contexte social, qui semble apparaître chez le bébé dès 9 mois". Le cerveau « copie sur son voisin »
« Cela m'a complètement déconcentré ! » Dans sa conférence d'après match à Roland-Garros, un joueur de tennis explique pourquoi son service a échoué dans le bas du filet sur la balle de match, après qu'un spectateur ait crié : « Double faute ! » Que s'est-il passé dans la tête de ce joueur ? Il suffisait pourtant d'une demi-seconde à son lobe frontal pour identifier que ce que venait de crier ce spectateur n'avait pas d'importance pour le jeu et pouvait être ignoré. Mais, comme dans le cas de la sonnerie de téléphone, son cerveau a surréagi en accordant beaucoup plus d'attention qu'il n'en fallait au stimulus distracteur. Le problème ne venait donc pas de la capture elle-même, mais du fait que la distraction s'était prolongée au-delà. Voilà ce que l'on appelle couramment « se laisser déconcentrer ».
La capture n'est qu'une première phase, très courte, pendant laquelle le cerveau oriente l'attention vers le distracteur pour l'identifier en une fraction de seconde. La raison pour laquelle les distracteurs ralentissaient si peu la recherche de la cible dans les expériences du chapitre précédent, c'est qu'elles n'induisaient que la capture de l'attention. Quand on distrait le sujet en faisant apparaître un A rouge pendant qu'il recherche un F rouge, ce distracteur n'a aucune des propriétés nécessaires pour captiver l'attention, car les propriétés qui contribuent à rendre un événement « captivant » sont très différentes de celles qui le rendent « capturant ». Dans des situations plus réalistes de la vie quotidienne, cette phase initiale de capture n'est que le début d'une cascade de processus moteurs, émotionnels et cognitifs qui permettent au cerveau de réagir à l'événement de façon adéquate.
Si, au moment de traverser la rue, j'entends un bruit de moteur sur ma droite, j'ai le réflexe de m'arrêter net et de me tourner pour regarder à droite. La capture de mon attention par le bruit de la voiture entraîne spontanément une réorientation de mes yeux, de ma tête et de mon buste, qui me permet d'identifier clairement l'origine de ce bruit. Il s'agit d'une prolongation de la capture oculomotrice que j'ai déjà évoquée. La capture de l'attention est donc aussi une capture du corps ; le monde extérieur tire sur notre corps comme un marionnettiste sur les fils d'une poupée.
Sur le plan neuronal, cette capture du corps fait intervenir le lobe pariétal – ce large continent cortical situé sous la tonsure des moines. Le lobe pariétal s'intéresse avant tout à l'espace. Il nous rappelle sans cesse l'existence d'une gauche et d'une droite, d'un en bas et d'un en haut, d'un près et d'un loin. C'est au sein du lobe pariétal que sont analysées la position, la trajectoire et la vitesse des objets qui nous entourent, le long de ce que l'on appelle la voie visuelle dorsale, surnommée « la voie où ? » (the where pathway). Cette voie dorsale remonte le long du « dos » du cerveau, à partir du cortex visuel primaire à la pointe arrière du cerveau jusqu'à son sommet, tout en haut du lobe pariétal près des régions motrices situées sous le sommet de la tête, dans le lobe frontal. Cette voie dorsale complète une autre voie visuelle, appelée voie ventrale, qui descend le long du lobe temporal — le pouce du gant de boxe — pour analyser les formes, les couleurs et la nature des objets situés devant moi ; les Anglo-Saxons l'appellent the what pathway — « la voie quoi ? ». Chaque objet, ou chaque être, qui se présente sous vos yeux génère donc deux vagues d'activation dans votre cerveau ; l'une descendant le long de la voie ventrale et l'autre remontant le long de la voie dorsale pour détecter, ensemble, qu'une Mercedes noire se dirige à vive allure dans votre direction.
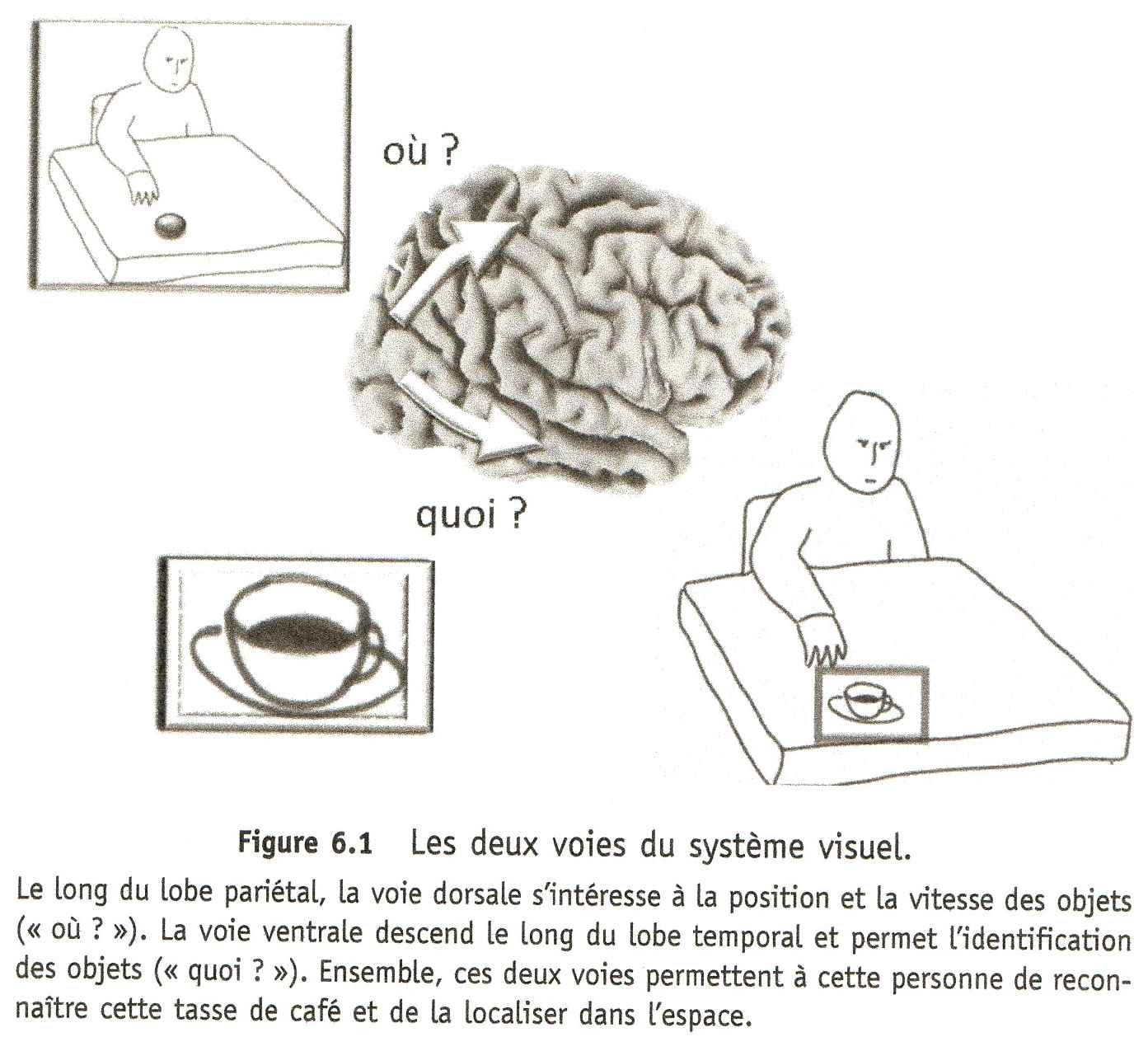
Imaginons que votre petite balade urbaine vous ait amené dans un café, pour y feuilleter tranquillement le journal. Vous êtes en train de parcourir les gros titres quand une forme claire attire votre attention vers la table d'à côté. Votre regard suit et vous identifiez un petit bol jaune rempli de cacahuètes. Sans réfléchir, vous en saisissez spontanément quelques-unes pour les avaler aussitôt, avant de retourner à votre lecture. Cette fois, le bol n'a pas seulement capturé votre attention et votre regard, mais également votre bras et votre main le long d'un comportement moteur assez complexe. Et comme vous n'avez repris votre tâche principale, la lecture du journal, qu'une fois la séquence d'action accomplie, cette phase de distraction s'est finalement prolongée jusqu'à l'ingestion des cacahuètes, bien au-delà de la simple capture de l'attention par le bol jaune.
Que s'est-il passé ? Pour attraper les cacahuètes, deux gestes sont nécessaires : un premier pour amener la main au niveau du bol et un second pour saisir les cacahuètes. La première opération ne pose aucun problème : en calculant la position du bol par rapport à la main droite, les neurones du lobe pariétal déterminent précisément le mouvement à réaliser pour l'atteindre. Il suffit ensuite que ces neurones communiquent cette information aux régions prémotrices pour que le mouvement soit préparé et réalisé. Pour saisir les cacahuètes, c'est une autre affaire, car le cerveau doit associer l'objet « cacahuètes » au geste particulier des doigts et de la main nécessaire pour piquer dans le bol.
Non content d'agir sur nos muscles, le monde extérieur sait aussi jouer sur nos émotions pour nous séduire et nous ensorceler. Nous savons déjà comment, grâce à l'amygdale notamment, les stimuli à forte valence émotionnelle capturent plus facilement l'attention que les stimuli neutres. Comme l'écrit le neurologue Antonio Damasio, « la réponse émotionnelle que nous ressentons en présence de tel ou tel objet joue un rôle majeur dans sa capacité à attirer ou non notre attention ». Mais ce n'est pas tout car, une fois l'attention capturée, la nature de ce que nous ressentons alors, au contact du distracteur, influe également sur la phase suivante : l'attention va-t-elle rester captive ou non ? Pour Damasio toujours, le rôle de l'émotion est à nouveau central : l'attention demeure ou non sur l'objet de la distraction en fonction de l'état émotionnel ressenti à son contact prolongé.
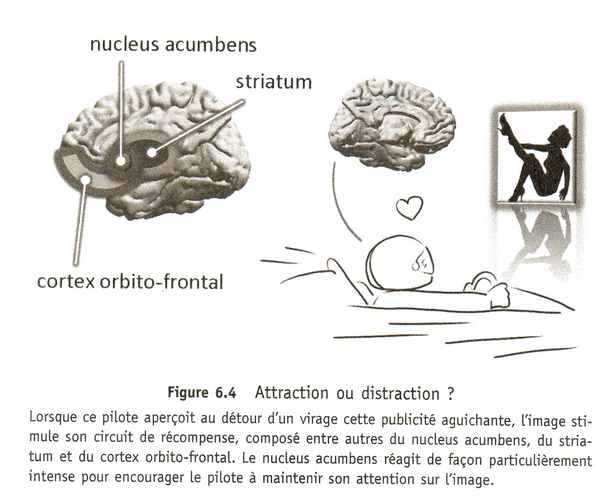
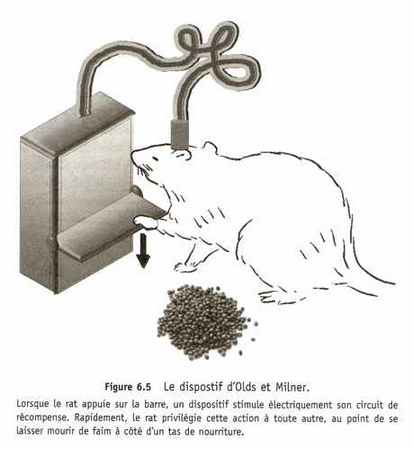
Pour l'heure, retenons simplement que le cerveau n'est pas un piano, ou alors un modèle étrange capable de poursuivre sa petite musique tout seul quand le musicien a cessé de jouer. Le cerveau est en permanence actif et les stimulations du monde extérieur ne font que moduler une activité déjà existante. Du point de vue du propriétaire de ce cerveau, le sujet de l'expérience, le petit concert prend principalement la forme d'images mentales, de sensations corporelles et d'impressions sonores, éventuellement verbales, accompagnées ou non d'un ressenti émotionnel ; tout ce que nous appelons couramment les pensées.
Ces études ont fait entrer les pensées spontanées dans le champ de l'investigation de la neuro-imagerie. Pour " faire scientifique ", le mot pensée est en passe d'être abandonné au profit d'un terme plus technique : activité cognitive indépendante des stimulations sensorielles (stimulus-independant cognition, ou SIC) ; et le thème y a gagné en respectabilité.
Le SIC correspond à un état dans lequel le cerveau semble se découpler de son environnement pour mener sa petite vie. Il s'oppose au SOC, ou stimulus-oriented cognition, état dans lequel l'activité cérébrale se soumet à la contrainte forte des stimulations du monde extérieur. En langage technique, l'étudiant qui part dans ses pensées pendant un cours bascule donc du mode SOC au mode SIC : SOC-SIC. Les études menées actuellement sur le SIC défrichent un champ de recherche très important concernant directement l'attention. Si vous souhaitez comprendre pourquoi vous décrochez parfois pendant que quelqu'un vous parle, c'est là qu'il faut chercher.
On s'attendrait logiquement à ce que le cerveau soit plus actif lorsqu'il doit faire quelque chose que lorsqu'il n'a rien à faire. Pourtant, la neuro-imagerie a révélé dès ses débuts, dans les années 1990, que certaines régions du cerveau sont moins actives quand une personne réalise une tâche que lorsqu'elle est censée ne rien faire. Comme on peut s'en douter, ce résultat a intrigué vivement les neuro-imageurs, qui ne savaient pas trop quoi faire de ces désactivations ni même quel sens leur donner. Mais la communauté neuroscientifique a fini par s'y intéresser sérieusement, sous l'impulsion notamment de l'Américain Marcus Raichle, quand plusieurs études ont révélé que les désactivations se produisaient toujours plus ou moins dans les mêmes régions du cerveau, quelle que soit la tâche demandée aux sujets. On a d'abord appelé ce réseau « réseau du repos », avant de le rebaptiser « réseau par défaut », bien conscient du fait que le cerveau n'est jamais vraiment au repos. Pour marquer davantage encore les esprits, Raichle parle maintenant de l'énergie noire du cerveau — « the brain's dark energy », en référence à la fameuse énergie noire de l'Univers. Quel que soit son nom, ce réseau joue un rôle important dans le processus de pensées spontanées, et les études menées par MacCrae, Gilbert et Schooler ont toutes révélé une activation du réseau par défaut pendant le SIC. Cela n'avait pas échappé à certains chercheurs spécialistes de ce réseau, qui avaient déjà rebaptisé l'état de repos REST, pour Random Episodic Silent Thinking (« Pensée silencieuse aléatoire et épisodique ») — excellent jeu de mots faisant référence à la traduction anglaise du mot « repos » : rest.
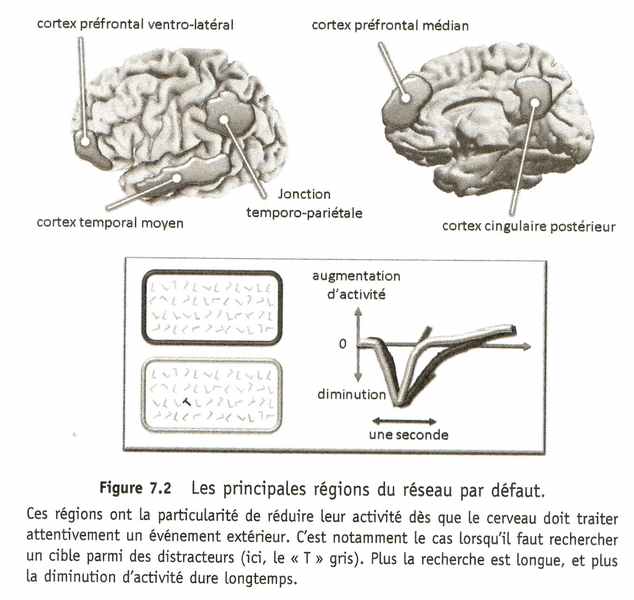
En termes simples, le système exécutif détermine quel comportement adopter et quels processus cognitifs mettre en jeu, face à telle ou telle situation.
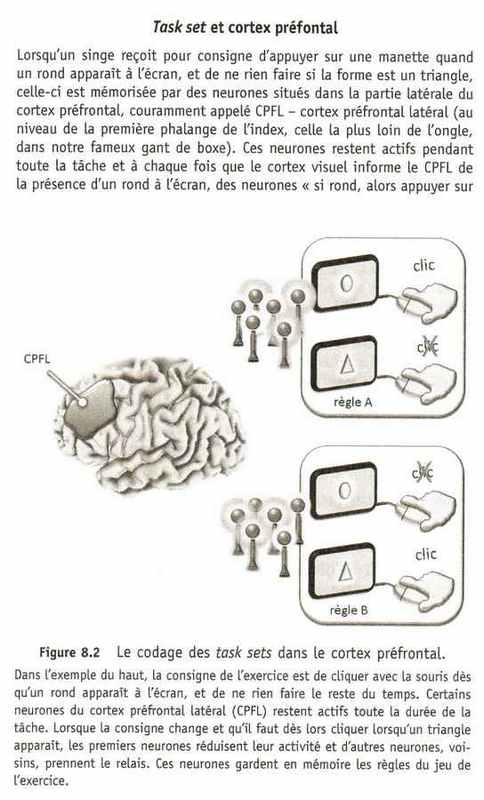
Le système exécutif permet donc d'associer de façon flexible un ensemble de stimuli avec un ensemble de processus moteurs ou cognitifs, selon un ensemble de règles de type « si A se produit, alors faire B » qui constitue Le task set. Cela ne signifie pas que toutes tes associations stimulus-réponse sont stockées dans le cortex préfrontal, loin de là. Avec l'apprentissage, les associations qui se répètent souvent finissent par avoir leur petit réseau de neurones spécialisé, et l'apparition du stimulus entraîne automatiquement la réponse qui lui est le plus couramment associée. Le cerveau prend l'habitude de réagir de cette façon-là à ce stimulus ; l'association est devenue habituelle. Le cortex préfrontal intervient surtout pour former des associations nouvelles, dont le cerveau n'a pas L'habitude. Dès qu'une association devient habituelle, et stéréotypée, elle est stockée dans d'autres structures, corticales comme le lobe pariétal – souvenez-vous du bol de cacahuètes – ou sous-corticales comme le Striatum. S'il n'y a pas de task set actif dans le cortex préfrontal, le cerveau peut tout de même réagir à l'apparition d'un stimulus, en fonction de ses habitudes, par automatisme. Le cas des pro-saccades est un bon exemple ; ce n'est qu'au moment où le cortex préfrontal rentre en action, avec l'activation d'un task set, que Le cerveau quitte le mode de pilotage automatique et passe dans un mode « contrôlé », pour réaliser des antisaccades par exemple. Contrôlé par quoi ? Par le système exécutif.
voir aussi : l'amygdale, amie de l'attention
Pourquoi les enfants désirent-ils tous le même jouet quand il en existe d’autres tout aussi attractifs, voire identiques dans la pièce ? Ce phénomène, connu de longue date, s’appelle en fait le désir mimétique. Il a été initialement décrit par un philosophe français nommé René Girard dans les années 1960. L’objet n’est pas voulu seulement pour ses qualités intrinsèques, ses performances, son utilité, sa physionomie, mais parce qu’il est convoité par d’autres. Il s’agit en quelque sorte d’une contagion motivationnelle d’un individu à l’autre pour l’obtenir. Plus les gens s’intéressent à lui, plus il devient désirable.
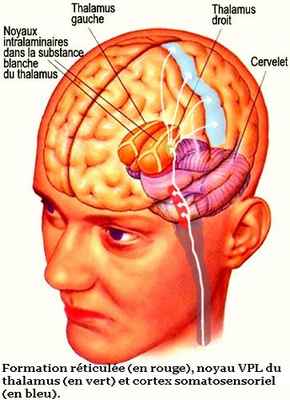
Le cortex somatosensoriel primaire (ou S1) reçoit les axones des neurones thalamiques du noyau ventral postérolatéral (ou VPL) tandis que le cortex somatosensoriel secondaire (ou S2) reçoit les informations nociceptives à la fois de l’aire S1 et des noyaux thalamiques. Si on assigne généralement à S2 un rôle dans la reconnaissance de la douleur et la mémoire des douleurs passées, S1 est associé pour sa part à la discrimination des différentes propriétés de la douleur.
L’organisation somatotopique est en effet préservée jusqu’au cortex S1 et rend possible la localisation de l’endroit douloureux sur le corps. Le degré d’activité des neurones de S1 correspond quant à lui à l’intensité du stimulus douloureux. Par exemple, plus l’eau qui coule sur votre main est chaude, plus vos neurones du cortex S1 vont déclencher d’influx nerveux. Les études d’imagerie cérébrale montrent que plus l’activité corticale dans S1 est intense, plus la douleur est évaluée comme intense subjectivement par le sujet.
D’autres études d’imagerie utilisant l’hypnose pour suggérer au sujet que l’intensité du stimulus douloureux est moindre que ce qu’elle est en réalité ont d’ailleurs démontré une diminution d’activité dans le cortex S1 dans ces conditions. Fait remarquable, si la suggestion hypnotique portait sur le caractère plus ou moins déplaisant du stimulus, l’activité dans le cortex S1 demeurait constante. Cependant, celle du cortex cingulaire antérieur, associée à la composante affective de la douleur, variait pour sa part en fonction de la suggestion, confirmant des rôles distincts pour ces deux régions.
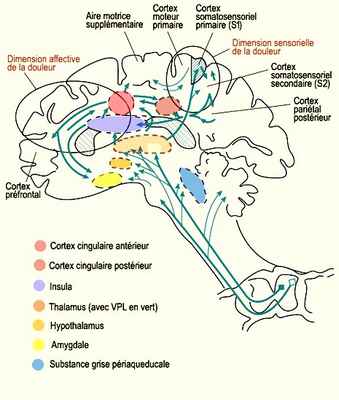
Cela dit, on admet aujourd’hui qu’il existe une spécialisation fonctionnelle au moins partielle des régions cérébrales impliquées dans les différentes composantes de la douleur. On tente donc maintenant d’associer à ces différentes composantes des sous-ensembles de structures cérébrales et de proposer ainsi un modèle fonctionnel global de la douleur. Étant donné la complexité du phénomène dont veulent rendre compte ces modèles, de vifs débats leur ont toujours été associés.
Grosso modo, on peut dire que la conception sous-jacente aux théories de la douleur est passée d'une causalité linéaire à une causalité circulaire.
Il distingue entre l’aspect somatique et mental des phénomènes affectifs liés à un objet émotionnellement pertinent : les émotions et les sentiments. D’un côté, les émotions sont définies comme aspect somatique et physiologique ; elles représentent l’ensemble de collection de réponses physiologiques induites par des objets émotionnellement compétents provenant de l’environnement ou du milieu interne de l’organisme. De l’autre, les sentiments sont conçus comme une sorte d’image mentale, subjectifs et privés.
La neurobiologie affirme que les émotions et les sentiments sont eux aussi le résultat du long cheminement de l'évolution animale, c'est bien cela ?
Au cours de recherches récentes, mon collègue David Rudrauf a étudié, par magnéto-encéphalographie, le cours du temps que suivent les émotions et les sentiments dans le cerveau humain. La magnéto-encéphalographie (MEG) est bien moins précise que la résonance magnétique fonctionnelle en termes de localisation spatiale de l'activité cérébrale, mais elle permet une remarquable estimation du temps que prennent certains processus dans des portions assez larges du cerveau. C'est précisément pour cette raison que nous l'utilisons dans ces recherches.
Au sein du cerveau, Rudrauf a suivi le temps mis par l'activité liée aux réactions émotionnelles et sentimentales face à des stimuli visuels plaisants ou déplaisants. Les chiffres parlent d'eux-mêmes. Du moment où les stimuli étaient traités dans les cortex visuels à celui où les sujets faisaient pour la première fois part de leur sentiment, près de 500 millisecondes s'écoulaient, soit environ une demi-seconde. Est-ce peu ? Est-ce beaucoup ? Tout dépend du point de vue. En « temps cerveau », c'est un intervalle énorme quand on songe qu'un neurone peut s'allumer en 5 millisecondes environ. En « temps d'esprit conscient », cependant, ce n'est pas beaucoup. Cela se situe entre les 200 millisecondes qu'il nous faut pour être conscients d'une structure perçue et les 7 à 800 dont nous avons besoin pour traiter un concept. Au-delà de ce niveau de 500 millisecondes, cependant, il faut noter que les sentiments peuvent persister de quelques secondes à quelques minutes et, bien évidemment, se réitérer en vertu d'une sorte de boucle réverbérante, en particulier s'ils sont importants.
Les tentatives menées pour décrire toute la gamme des émotions humaines ou pour les classer ne sont pas particulièrement intéressantes. Les critères utilisés par les classifications traditionnelles sont entachés de défauts et tout tableau peut être critiqué au motif qu'il n'en inclut pas certains et accorde trop d'importance à d'autres. À vue de nez, on doit réserver le terme émotion pour désigner un programme assez complexe d'actions (qui doit comporter plus d'une ou deux réponses de type réflexe) qui est déclenché par un objet ou un événement identifiable, un stimulus émotionnellement compétent. On peut estimer que ce qu'on appelle les émotions universelles (peur, colère, tristesse, joie, dégoût et surprise) satisfont ces critères, même si j'ai mis du temps à admettre la présence de la surprise dans cette liste. Ces émotions sont à coup sûr produites dans toutes les cultures et elles sont faciles à reconnaître car une partie de leur programme d'action — les expressions du visage — sont très caractéristiques. Elles sont présentes même dans des cultures dépourvues des noms distinctifs pour les désigner. On doit à Charles Darwin d'avoir le premier reconnu cette universalité non seulement chez les êtres humains, mais aussi chez les animaux.
À côté des émotions universelles, on trouve deux groupes bien identifiés d'émotions qui méritent une mention spéciale. Il y a quelques années, j'ai attiré l'attention sur l'un d'eux et je lui ai donné un nom – émotions d'arrière-plan. Ce sont par exemple l'enthousiasme et le découragement, qui peuvent être déclenchés par diverses circonstances factuelles de la vie, mais aussi par des états intérieurs comme la maladie et la fatigue. Encore plus que pour les autres émotions, le stimulus émotionnellement compétent des émotions d'arrière-plan peut opérer en coulisse : il déclenche l'émotion sans qu'on ait conscience de sa présence. Réfléchir à une situation qui s'est déjà produite ou en envisager une qui est possible peut déclencher de telles émotions. Les émotions d'arrière-plan sont, très proches des sentiments primordiaux, mais elles en diffèrent par leur profil temporel plus circonscrit et leur identification plus nette d'un stimulus.
Exprimer ses émotions est parfois difficile pour soi, mais aussi pour les autres pas toujours prêts à les recevoir.
Parler aide à se sentir bien, partager ses émotions favorise la communication et crée du lien. Les personnes se soutiennent, s’apprécient ou se respectent davantage.
Evoquer ce qu’on ressent, en parler à ses proches, partager autour de soi le monde intérieur de ses émotions voilà qui rend la vie plus facile, qui améliore le lien social et la santé. Créer, fabriquer, dessiner, inventer… est également une source d’expression accessible à tous.
Identifier une émotion permet de ne pas simplement la subir.
En observant ses pensées, ses tendances à l’action et son corps on apprend à reconnaître et à identifier son état émotionnel.
La compréhension des émotions des autres implique de les reproduire comme dans un miroir à l’intérieur de soi. Identifier ses propres émotions aide à reconnaître celles des autres.
Transformer son émotion demande de s’y confronter, de faire des compromis pas tant pour la réduire que pour en faire quelque chose.
Ce réel travail sur soi peut se mener de différentes façons.
Les psychomotriciens proposent des techniques de relaxation par la respiration (la première des nécessités, c’est de respirer), par le recours à des images mentales ou des positions du corps. Il s’agit de réfléchir à se détendre.
Tester la réévaluation cognitive, cette méthode demande de comprendre que la cause de notre affect n’est pas liée à une situation mais à l’évaluation que nous en faisons. Il faut chercher une autre façon de considérer les choses, de les remettre en perspective. Souvent il s’agit d’adopter le point de vue d’autrui pour aboutir à un ressenti émotionnel différent.
Il peut s’agir aussi de rechercher un contact social accru malgré les tendances au repliement.
Pour renforcer et prolonger les émotions positives, demander au corps : exprimer au maximum, par les gestes, la parole, les expressions du visage… Cette théorie de régulation physique repose sur la perception intime des émotions qui s’enracine dans l’expression du corps.
Enfin, les activités manuelles permettent de libérer l’imagination avec les émotions, et inversement !
Les personnes se comportant comme animées de sentiments positifs peuvent sentir apparaître une vraie émotion positive ou encore attirer l’attention des autres et leurs réactions positives.
Comprendre aide à vivre avec ses émotions.
Les émotions sont liées à des besoins qui sont ou ne sont pas satisfaits. Lorsqu’une explosion d’émotion survient il ne faut pas forcément l’attribuer à un déclencheur trop proche ou trop évident mais s’interroger sur les causes profondes de cette manifestation.
Bénéficier de ses émotions, en profiter pour créer quelque chose, prendre de meilleures décisions et ainsi s’aider à vivre mieux et être heureux sur le plan professionnel, personnel et dans ses relations à l’autre en général.
Les émotions peuvent déformer notre réalité du moment, mais nous alertent bien souvent sur notre état profond.
Ils n’ont pas le langage mais communiquent déjà. Ils expriment des affects, des émotions qui sont attachés à des besoins : manger, dormir…
Les bébés ressentent très tôt des sensations, du bien être et de la douleur, des plaisirs et des déplaisirs. Leur entourage participe largement à leur définition, leur compréhension et à l’expression de leurs émotions.
L’enfant vit entièrement ses émotions sans arrière-pensée, sans pouvoir les refreiner ni les raisonner.
Les enfants croient leurs émotions éternelles, infinies.
L’instant présent dure toute la vie pour eux.
Le monde émotionnel des enfants est très différent de celui des adultes qui est rationnalisé et d’une certaine façon désenchanté. Le cortex préfrontal teste chez l’adulte si ce que nous voyons est extérieur à nous ou le produit de notre psychisme. Chez l’enfant, il y a une maturité d’enfant et du coup tout est lié, le réel et l’imaginaire.
L’enfant est guidé par ses proches, son environnement, sa culture.
En ne cédant pas sur le champ aux moindres désirs des enfants on stimule le fonctionnement de la partie frontale du cerveau qui peut faire patienter les zones plus profondément enfouies du plaisir. Parents, il faut doser la frustration pour aider son enfant à devenir un être mûr émotionnellement non dominé par la tyrannie de ses émotions.
Certaines de ses émotions sont perçues comme négatives.
Les colères des enfants sont violentes et prennent la forme du caprice. Lorsqu’ils ne peuvent avoir un plaisir qu’ils ont anticipé leur cerveau cesse de fabriquer dopamine et enképhalines ce qui entraîne une réaction réflexe d’agression liée à la frustration.
L’immaturité des réseaux entre les zones du plaisir, les zones de l’agression et les zones frontales du cortex ne lui permet pas de maitriser ses réactions. Ses réactions sont alors impulsives. Chez l’adulte les zones frontales prennent en compte les contraintes de l’environnement et invitent à patienter, le plaisir viendra bientôt.
L’enfant a du mal à contenir sa tristesse. Là encore les connexions entre les différentes zones de son cerveau qui permettent de contenir ses émotions ne sont pas tout à fait en place. Il ne fera que gagner en stabilité émotionnelle grâce au développement du cortex préfrontal, le système cérébral de la réévaluation des émotions.
Le cerveau subit de profondes mutations à l’adolescence. Les neurones changent, la circulation de l’information est bien plus rapide, multipliant les capacités intellectuelles.
En parallèle le nombre de récepteurs à la dopamine diminue, les ados ont donc besoin de stimulations plus intenses pour ressentir la même émotion que dans l’enfance.
Le caractère rebelle s’explique par des zones non encore matures qui permettent de prévoir les conséquences de ses actes, notamment les connexions entre le cortex préfrontal et l’amygdale.
Le cerveau a bien des maturités différentes selon les âges.
Toutefois la création et un vocabulaire émotionnel enrichi sont un rempart aux conduites impulsives et agressives car il devient plus facile d’exprimer son émotion et pour les autres de la comprendre. Mettre en mot le ressenti émotionnel permet de ne pas y céder immédiatement, muscle le cerveau en créant des connexions utiles pour contrôler ses élans.
Le passage à l’âge adulte n’est pas chose facile. L’adolescent doit être guidé et responsabilisé dans le même temps. Le désinvestissement des parents et la disparition des rites de passage entraînent une maturation plus difficile des zones cérébrales qui contrôlent les émotions.
..c'est un état second dans lequel on ne s’obéit plus.
Elle décuple les forces, déforme le visage, altère la voix.
C’est un programme comportemental automatique devant lequel la raison s’efface.
Elle perturbe le jugement. Le champ de réflexion se restreint et les diverses solutions d’un problème se réduisent souvent à une solution unique une simplification qui offre parfois un surcroît d’efficacité dans des situations extrêmes. Elle peut aussi laisser des traces profondes dans les relations que l’on entretient avec les autres.
En colère, on a besoin d’aide, besoin d’être entendu. C’est une réaction de défense ou une façon de décharger un trop plein de frustration.
La noradrénaline accélère le rythme du cœur
Le cortisol mobilise l’énergie en produisant un afflux de sang dans les muscles
La testostérone apporte un surcroît d’agressivité.
La déformation de la bouche découvre les dents, une technique ancienne et efficace d’intimidation.
Le plissement du front et le rehaussement des paupières en réduisent l’ouverture ce qui protège les yeux en situation de combat.
Le visage pâlit, le cortisol produit un afflux de sang dans les muscles, du même coup le sang quitte le visage.
A ce même titre, le langage offre une autre alternative, la dispute verbale.
A trop contenir sa colère le corps est dans un état de tension permanent qui peut se traduire à long terme par des problèmes cardio-vasculaire et une baisse de l’immunité.
Il faut l’admettre sans la masquer. Celui qui accepte sa colère peut encore changer le cours des choses, celui qui l’occulte a renoncé à les changer.
L’ennui aussi est une émotion !
Il est lié à un certain type d’expériences avec ses manifestations corporelles propres.
Il se manifeste dans le corps par une baisse de l’énergie et une somnolence envahissante, d’une torpeur perçue comme désagréable. Les bâillements sont plus ou moins faciles à réprimer. Le visage prend une expression typique de l’ennui.
Biologiquement la chose la plus rationnelle à faire face à l’ennui, c’est se reposer.
L’origine de l’ennui est souvent jugée négativement.
Eviter de s’ennuyer est considéré comme un impératif dans notre société. Nous n’aimons pas nous ennuyer mais avons-nous raison de lutter contre l’ennui ?
L’ennui possède une fonction naturelle.
Il peut être causé par des objets qui ne nous semblent pas intéressants dans le contexte où ils se présentent. Le cerveau humain dispose de capacités cognitives limitées. Il ne peut mobiliser son attention sur un très grand nombre de stimuli au même moment et accomplir de nombreuses tâches en parallèle. Au même titre que l’oubli, l’ennui préserve le cerveau de la « surchauffe ». C’est là sa fonction biologique, sélectionner les stimuli.
Détecter très vite les stimuli les plus importants, les plus surprenants dans un contexte cognitif donné permet de fournir des informations nouvelles au cerveau.
Les phénomènes répétitifs sont eux ennuyeux à observer comme les tâches cognitives complexes en raison de leur difficulté.
L’ennui peut aussi être sans objet, surgit tout droit de notre propre intériorité. Dans cette expérience de vide et de détachement vis à vis des choses, l’homme saisit son essence, sa vraie nature. Il fait l’expérience du temps qui passe ou qui refuse de passer.
Notre attitude naturelle est de laisser passer le temps, de laisser l’ennui s’endormir. La posture philosophique invite à prendre de la distance vis à vis de l’attitude naturelle. Il s’agit donc de laisser l’ennui être éveillé parfois pour aller à la rencontre de soi…
L’ennui dans les apprentissages
Pour apprendre ou apprendre à apprendre il faut parfois accepter de s’ennuyer, de ne pas être systématiquement excité intellectuellement par une activité ou un exercice.
Les élèves ne sont pas naturellement aptes à évaluer l’intérêt d’un sujet. Une bonne pédagogie devrait donc reposer sur la recherche de moyens pour rendre plus attractifs les contenus enseignés mais aussi sur la création d’un lien de confiance avec l’enseignant. L’ennui doit être apprivoisé et chercher à l’éliminer n’est pas nécessairement souhaitable.
La plupart des scientifiques pensent que les humeurs apparaissent essentiellement pour des raisons que celui qui les vit ne comprend pas.
Elles peuvent être générées par des changements neuro hormonaux qui ne sont pas directement liés à ce qui survient autour de nous. Toutefois certains évènements peuvent déclencher une humeur ; par exemple, si vous manquez de sommeil, vous risquez davantage d’être irritable ou frivole ou de vous moquer des choses dont vous ne vous moqueriez pas habituellement.
Chaque humeur est saturée d’une émotion particulière.
Donc, quand une personne est d’humeur irritable, la déformation de la réalité induite sert à favoriser la colère. Elle cherche une occasion de se fâcher. Mais souvent on ne se rend même pas compte qu’on est d’humeur irritable.
C’est plus qu’un penchant : vous êtes biologiquement prêt à vous fâcher. L’humeur fait que vous déformez la réalité pour qu’elle lui corresponde. Mais, si une personne est plus à l’écoute d’elle-même, elle comprendra qu’elle est dans un état irritable, et qu’elle ferait mieux de ne pas s’emporter.
Etre impatient, c’est avoir du mal à supporter quelque chose, quelqu’un ; une incapacité à se contraindre ou à attendre. Cette difficulté est plus grande encore quand entre en jeu un contexte émotionnel particulier ou l’envie de satisfaire un besoin. Cette incapacité à supporter une attente, voire une frustration, peut provenir de la personnalité, de la situation vécue, de l’éducation ou de la culture.
Nous sommes tous plus ou moins «extraverti» (tendance à ressentir des émotions positives, à agir, à rechercher des stimulations externes ainsi que la compagnie d’autrui), «névrotique» (tendance à ressentir plus souvent des émotions négatives), avec un «caractère agréable» (tendance à montrer de la compassion ainsi qu’à coopérer au lieu d’avoir une attitude suspicieuse et opposante envers autrui), un «caractère consciencieux» (tendance à s’imposer une discipline propre, à agir en conséquence de ses responsabilités, de ses devoirs afin d’atteindre ses objectifs) ou une «ouverture à l’expérience» (propension générale pour l’art, l’émotion, l’aventure, les idées originales, l’imagination, la curiosité).
Derrière les cinq traits généraux de la personnalité de ce modèle apparaît la notion d’impulsivité.
L’impulsivité conduit vers des paroles, des comportements exprimés de manière prématurée, immédiate (sans réflexion ou contrôle). L’impulsivité est aussi liée à une difficulté à reporter ou à attendre une gratification ou une récompense. Elle est renforcée plus encore dans des contextes émotionnels.
Bien que le concept d’impulsivité soit différent de celui d’impatience, les attentes, la manière de fonctionner d’un individu très impulsif favoriseront une attitude impatiente et ce, plus particulièrement dans des contextes délicats.
Ces manifestations sont présentes à une moindre intensité chez tout un chacun.
L’intensité de l’impatience ressentie ou exprimée dépend en partie de la situation dans laquelle nous nous trouvons et de l’importance que revêt cette même situation à nos yeux.
De même, cette impatience peut se manifester plus particulièrement dans des situations coutumières. Dans des contextes habituels pour l’impatient, une action peut paraître évidente à effectuer pour ce dernier, cependant ce même acte pourrait être ardu à réaliser pour une personne de l’entourage; ces situations sont donc propices à un manque de tolérance, car la personne de l’entourage tardera à réaliser l’action attendue.
Cette même impatience pourrait également se manifester lors d’une divergence dans la méthode à appliquer, l’un des deux individus estimant que sa méthode est la meilleure et ainsi ne laissant pas le temps à l’autre de comprendre ses erreurs ou d’appliquer une méthode peut-être aussi ou plus efficace.
Selon le style éducatif des parents, les enfants ont plus ou moins été habitués à la frustration. Certains parents en limitant l’accès aux sucreries, à des cadeaux constants ou à la satisfaction immédiate de tous désirs ont ainsi appris à leur enfant à reporter une envie, un désir. Bien évidemment, cet apprentissage est plus ou moins facile selon l’enfant et son tempérament. Néanmoins, cette capacité à reporter un désir, une réponse immédiate peut favoriser un comportement patient et compréhensif, l’équilibre.
Il est très mal vu dans certaines cultures de perdre son sang froid ou de se mettre
en colère à un guichet par manque de patience ; selon notre culture, certains comportements sont plus ou moins valorisés ou tolérés.
Enfin, le développement de la technologie et le rapport que nous entretenons avec elle favorise notre impatience. On appuie sur un bouton, on touche un écran et tout arrive dans l’instant.
Ces conditions sont un terreau formidable pour le développement de l’impatience. Et quand nous traversons une crise émotionnelle ou personnelle, elle fleurit.
L’impatience induit toutes sortes de comportements négatifs: incivilités, insultes, bagarres.
Ces réactions s’avèrent dommageables pour les autres mais aussi pour soi.
...c’est du plaisir mais plus encore, une sensation enivrante et sereine, un sentiment de bien-être. Le corps se détend, relâché et éveillé. La sensation est intense d’être là, pris tout entier, plein d’une énergie dure à contenir.
Le cerveau libère des hormones relaxantes et excitantes en même temps : la dopamine (hormone du plaisir), la sérotonine (hormone de la bonne humeur), l’adrénaline (hormone de l’excitation), l’ocytocine (hormone du lien social).
La dopamine irrigue un groupe de neurones, situé au cœur du cerveau entre l’aire tegmentale ventrale et le noyau accumbens, qui tient un rôle particulièrement important dans le circuit de la récompense. Ce système de récompense est un système fonctionnel fondamental, indispensable à la survie, car il fournit la motivation nécessaire à la réalisation d'actions ou de comportements adaptés. Chef d’orchestre d’un ballet chimique, il organise les informations reçues d’autres régions périphériques (l’amygdale, le septum, le cortex préfrontal…), leur attribue à toute vitesse des valeurs de plaisir ou de déplaisir et enclenche les mécanismes de réaction. L’hypothalamus prend le relais pour préparer notre corps en envoyant des messages chimiques. Ce système dopaminergique plus on y est exposé, plus on est motivé pour en retrouver la saveur. Il est la source du plaisir et de la motivation qu’on ressent dans la joie.
Mais dans la joie au contraire du simple plaisir l’insula et le cortex somato-sensoriel vont s’activer.
Le cortex somato-sensoriel reçoit les informations sensorielles en provenance du corps. Sous l’effet de la joie, il est plongé dans un climat de plaisir et de bien-être ce qui provoque un ressenti tout particulier de notre propre corps.
L’insula, quant à elle, donne le sentiment d’être pris dans tout son être.
L’expression principale de la joie, c’est le rire. Rire a une fonction sociale, alerter les membres d’un groupe. C’est un signal sonore reconnu par les autres comme une absence de danger.
La joie est un antidote à la maladie et une garantie de longévité. Pour vivre vieux en bonne santé vivez joyeux. Elle stoppe la libération de cortisol (hormone du stress) et renforce les défenses naturelles.
La joie est une des émotions les plus profitables à l’être humain. Elle permet de prendre de la hauteur, de gagner en justesse d’analyse. L’émotion positive modifie notre façon de réfléchir, elle nous permet de prendre du recul sur le réel.
La joie se manifeste en l’absence de menace. L’esprit peut alors explorer de nouvelles pistes.
C'était mieux avant !
Nous sommes tous sujets à la nostalgie. Enfants ou adultes, on se remémore tous avec nostalgie des événements du passé.
On y associe généralement des mots positifs, « chaud au cœur », « bon vieux temps » ou encore « enfance ». Elle se développe avec l’âge.
Le narrateur joue le rôle principal. La nostalgie est une forme particulière de mémoire autobiographique. Les bouffées de nostalgie concernent des relations humaines passées, riches et empreintes de sentiments, d’honnêteté et de chaleur. De telles réminiscences sont un refuge où il fait bon se blottir dans les moments difficiles.
On se souvient de bonnes choses qui apportent du réconfort et qui nous représentent des personnes ayant compté pour nous. Évidemment, la tendance à donner plus d’importance aux événements positifs et moins aux événements négatifs est une loi fondamentale de la psychologie.
La nostalgie ne s’embarrasse pas des faits et de l’analyse objective de la réalité. La mémoire gomme les évènements négatifs. Les personnes effacent de leur cerveau ce qui ne leur plait pas pour ne retenir que le meilleur d’un passé devenu sans reproches.
On devient nostalgique lorsqu’on se sent triste ou seul. La fonction réparatrice de la nostalgie semble plus grande encore dans les situations d’abandon.
Le fait de se plonger dans ses souvenirs améliorerait l’humeur, augmenterait l’estime de soi et renforcerait les relations avec autrui.
Elle aurait pour fonction de restaurer, par sa charge affective positive, une humeur temporairement assombrie.
Il suffit parfois d’évoquer son propre passé, pour faire surgir une meilleure image de soi-même. La nostalgie fait du bien. Les élans de nostalgie nous donnent l’impression d’être plus aptes socialement, et, dès lors, de moins craindre l’isolement.
Les nostalgiques sont bien plus confiants que d’autres dans le soutien que peut leur apporter leur entourage en cas de coup dur. Il s’agit d’un effet sans doute inhérent à la nature humaine : la capacité que l’on a d’évoquer de belles images du passé rend plus confiant en l’avenir…
La nostalgie, c’est un élixir d’optimisme.
… est universelle. Elle fait partie du registre émotionnel de base de l’humanité. C’est une émotion parmi les plus primitives et les plus puissantes. Ses ingrédients s’enracinent dans un passé ancestral peuplé d’animaux inquiétants et de milieux hostiles. Elle s’appuie sur ce qu’on ne voit pas (le noir) et ce qu’on ne comprend pas : l’inconnu ou l’irrationnel.
La peur se nourrit d’elle-même, elle s’amplifie jusqu’à la panique.
La peur peut se traduire par trois types de comportement, trois réactions de survie : l’immobilisation, la fuite ou la lutte.
La peur la mieux connue des enfants est la peur du noir.
Quand la lumière s’éteint et que l’on se retrouve seul, l’angoisse monte : un loup sous le lit, un monstre dans le placard. Les centres nerveux qui suscitent la peur sont déjà matures dans le cerveau de l’enfant, alors que ceux qui la tempèrent ne le sont pas. Pour se rassurer il n’a que la vue et s’il ne voit rien…
L'information sur les stimuli externes arrive à l'amygdale par une voie directe provenant du thalamus (la voie basse) ou par une voie passant de plus par le cortex. La voie basse est plus courte et donc plus rapide que celle arrivant du cortex. Mais comme elle court-circuite le cortex, elle ne peut bénéficier du traitement cortical et ne fournit donc à l'amygdale qu'une représentation grossière du stimulus. C'est une voie rapide et grossière de traitement (Chez le rat il faut 12 millisecondes "12 millièmes de seconde" pour qu'un stimulus arrive à l'amygdale par le thalamus et près de deux fois plus s'il doit passer par la voie corticale). La voie directe nous permet de commencer à répondre à des stimuli potentiellement dangereux avant de savoir pleinement de quoi il s'agit. Cela peut être très utile dans des situations dangereuses. Cette voie directe pourrait être à l'origine de réponses émotionnelles que nous ne comprenons pas, celles qui se produisent probablement chez chacun d'entre nous de temps en temps mais pourraient représenter un mode de fonctionnement prédominant chez les individus atteints de certains troubles émotionnels.
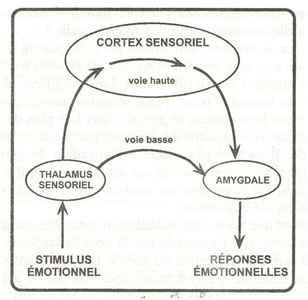
Le noyau latéral de l'amygdale est considéré comme la région de l'amygdale qui reçoit les entrées des stimuli (auditif ou visuel) et sa région centrale comme celle des réponses à ces stimuli.
En présence d'un danger ou de stimuli qui l'annoncent, on observe des réponses comportementales, végétatives ou endocrine et la modulation de réflexes. Chacune de ces réponses est régulée par des groupes différents de signaux issus du noyau central de l'amygdale. Des lésions de ce noyau bloquent l'expression de toutes ces réponses, tandis que celles effectuées sur leur voie spécifique les suppriment individuellement. On peut voir ici quelques signaux fournis par l'amygdale centrale. Abréviations : SG, substance grise centrale ; HL, hypothalamus latéral ; NPV, noyau paraventriculaire de l'hypothalamus (qui reçoit directement les signaux de l'amygdale centrale et par le truchement du noyau du lit de la strie terminale) : RPC, reticulopontis caudalis.
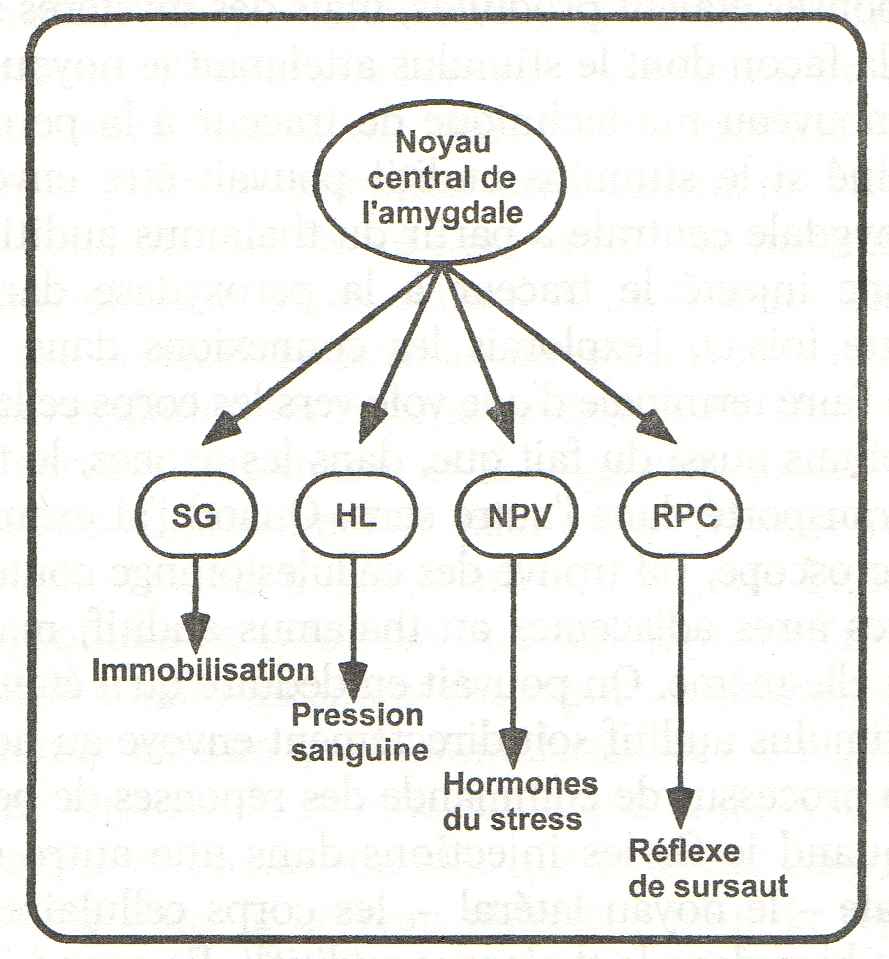
L'amygdale reçoit des signaux issus d'étapes très variées du traitement cognitif. Grâce à ceux provenant des aires sensorielles thalamiques, les fonctions émotionnelles de l'amygdale peuvent être déclenchées par de faibles stimuli, tandis que les signaux du cortex sensoriel permettent à des niveaux plus complexes du traitement des stimuli (objets, événements) d'activer l'amygdale. Les signaux issus de l'hippocampe jouent un rôle important en situant le contexte émotionnel. La connaissance des aires corticales qui se projettent sur l'amygdale et de leur fonction peut permettre de prévoir de quelle manière elles peuvent participer aux réactions de peur. L'anatomie peut ainsi donner un nouvel éclairage à la psychologie.
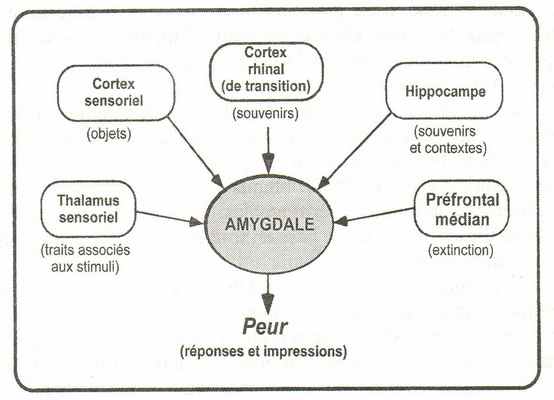
Il n'est pas déraisonnable de penser qu'en connaissant les différents signaux arrivant à l'amygdale de différentes aires que jouent ces dernières et en ayant une idée de la fonction dans la cognition, on puisse faire des hypothèses valables sur les types de représentations cognitives capables de susciter des réponses de peur. Et par la même occasion, si nous savons comment le cerveau réalise certaines fonctions cognitives et que nous pouvons déterminer comment les régions cérébrales effectuant ces tâches sont en rapport avec l'amygdale, nous pouvons peut-être arriver à une idée plausible de la manière dont ce type de cognition peut susciter la peur.
Vous marchez dans la rue et vous remarquez que quelqu'un court vers vous. La personne, en vous rejoignant, vous frappe à la tête et vous vole votre portefeuille ou votre sac à main. La prochaine fois que l'on courra vers vous, il y a des chances qu'un ensemble de réactions typiques de la peur se déclenchera. Probablement vous vous immobiliserez et vous vous préparerez à vous défendre, votre pression sanguine et votre rythme cardiaque augmenteront, vos paumes transpireront, les hormones de stress commenceront à circuler dans votre sang, etc. La vue de quelqu'un courant vers vous est devenue un stimulus conditionnel de peur. Mais supposez que vous vous retrouviez plus tard dans la rue où vous avez été agressé(e). Bien que personne ne coure vers vous, votre organisme va encore engager ses réactions de défense. La raison en est que vous n'avez pas été conditionné seulement par le stimulus immédiat qui a été directement associé au traumatisme (la vue du voyou courant vers vous) mais aussi par les autres stimuli qui se sont présentés juste à ce moment-là. Ils forment le contexte dans lequel s'est déroulée l'agression et, comme la vue de l'agresseur, ils ont aussi été conditionnés par l'expérience traumatique.
Le conditionnement par le contexte est en un sens un apprentissage fortuit. Au cours du conditionnement, le sujet porte son attention sur le stimulus le plus évident mais les autres stimuli sont enregistrés par la même occasion. C'est d'une grande utilité du point de vue évolutif. Notre lapin qui échappe au renard se trouve conditionné non seulement par les stimuli directement et immédiatement en rapport avec l'arrivée de son prédateur - sa vue, son odeur, et les sons qu'il produit en attaquant - mais aussi par l'endroit où la rencontre a eu lieu, le point d'eau et ses alentours. Ces stimuli supplémentaires sont très utiles pour étendre l'impact du conditionnement au-delà des stimuli les plus évidents et directs, ce qui permet à l'organisme d'avoir recours à des indices, même lointains, pour éviter le danger ou y échapper.
Ce qui est intéressant dans un contexte, c'est qu'il ne correspond pas à un stimulus en particulier mais à un ensemble de stimuli. On a cru pendant un moment que l'intégration des stimuli individuels pour former un contexte global était une fonction de l'hippocampe. Cette région, à la différence de l'amygdale, ne reçoit pas d'informations des régions du cerveau qui traitent les stimuli sensoriels individuels tels que la lumière ou le son. Ces éléments propres à un lieu sont regroupés avant d'atteindre l'hippocampe, et l'une des tâches de ce dernier est de créer une représentation du contexte contenant non pas les stimuli pris un à un mais les relations qui les unissent.
Le promeneur marchant dans la forêt se retrouve soudain devant un serpent enroulé derrière un tronc d'arbre (en haut à droite). Le stimulus visuel est d'abord traité par le thalamus dans le cerveau. Une partie du thalamus transmet l'information, presque archétypale, à l'amygdale. Cette transmission rapide et grossière permet au cerveau de commencer à répondre au danger possible représenté par un objet fin, incurvé qui peut être un serpent, un bâton ou un autre objet anodin. En même temps, une autre partie du thalamus envoie aussi l'information visuelle au cortex visuel (elle code mieux les détails du stimulus que celle envoyant ses signaux à l'amygdale). Le cortex visuel se met alors à créer une représentation précise et détaillée du stimulus. Les signaux issus de ce traitement sont également envoyés à l'amygdale. Bien que la voie corticale fournisse à l'amygdale une réprésentation plus détaillée que la voie directe thalamus-amygdale, elle met plus de temps à parvenir au cortex. En cas de danger, il est très utile de pouvoir répondre rapidement. Le temps gagné pour mettre en branle l'amygdale peut être une question de vie ou de mort. Il vaut mieux avoir traité un bâton comme un serpent que de ne pas avoir réagi à un possible serpent. L'essentiel de ce que nous savons sur ces voies a en fait été appris par des études du système auditif et non visuel, mais les principes d'organisation semblent s'appliquer de la même manière.
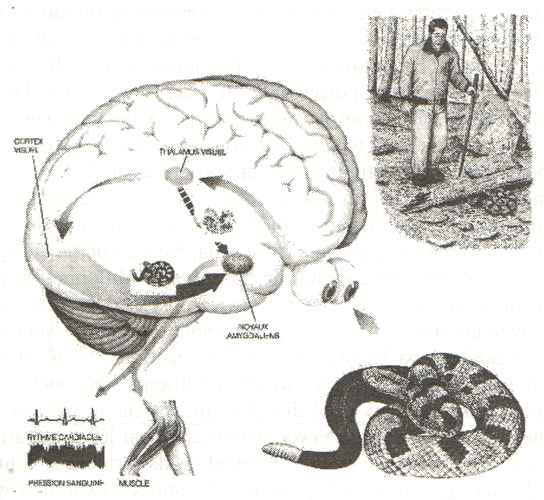
Quand la peur prend les commandes-1
Quand la peur prend les commandes-2
Quand la peur prend les commandes-3
1 - extraction :
Le cerveau semble extraire l'information en provenance du monde extérieur en deux temps. Dans le cas de la vision, par exemple, l'information transite d'abord selon une voie ascendante — en anglais bottom-up, du bas vers le haut :
depuis les premiers étages du cortex visuel, dans les aires dites primaires situées à l'arrière du cerveau, jusqu'aux étages les plus élevés, situés principalement à l'avant du cerveau dans le lobe frontal.
Cette propagation de l'information est aussi qualifiée de feedforward, qui signifie « vers l'avant ».
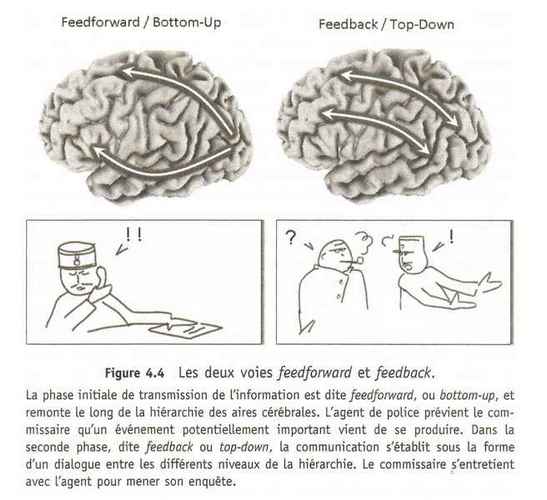
Évidemment, toute la question est de savoir quels sont les critères qui décident de la mise en action du lobe frontal. Ces critères prennent d'abord en compte les caractéristiques physiques simples des stimuli, comme leur taille ou leur luminosité. Le flash d'un appareil photo dans le noir attire naturellement l'attention. Mais il existe d'autres critères plus complexes dont les neurosciences cognitives peinent à faire l'inventaire. La capture de l'attention par son propre prénom est un bon exemple, mais il y en a beaucoup d'autres. Nous nous sommes par exemple aperçus dans l'expérience des histoires que les sujets n'ignoraient pas systématiquement tous les mots rouges. Certains d'entre eux parvenaient à se faufiler jusqu'au lobe frontal, même sans leur petit badge vert les y autorisant. Il s'agissait souvent de noms propres connus, comme « Manhattan » ou « Paris » ; les mots plus ordinaires, comme « faire » ou « plus » étant apparemment plus faciles à ignorer. Les sujets avaient également plus de mal à ignorer les mots rouges s'insérant parfaitement dans la structure de la phrase verte en train d'être lue. Une personne venant de lire les trois mots verts suivants : « le — petit — chat », avait plus de mal à ignorer le mot « mange » que le mot « maison », parce que la phrase « le petit chat mange » a un sens, alors que « le petit chat maison » n'en a pas.
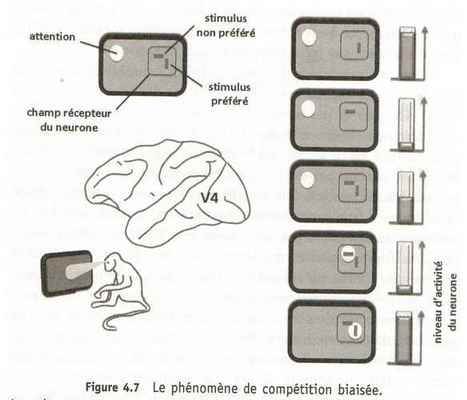
Dans le cas de la plus fréquente des démences, celle de type Alzheimer, la dégénérescence neuronale à l’origine des déficits se déploie de façon séquentielle caractéristique dans le cerveau. De sorte que les symptômes de l’Alzheimer apparaissent dans un ordre qui, sans être complètement prévisible, est typique de cette démence.
On utilise généralement les termes « léger, modéré et avancé » pour définir trois grands stades de l’évolution de la démence de type Alzheimer. Évidemment, les transitions entre ces différents stades ne sont pas coupées au couteau, pas plus que chaque patient va expérimenter les symptômes dans le même ordre, avec la même gravité ou la même durée (voir l’encadré ci-bas sur l'espérance de vie).
Il n’empêche que les associations de personnes atteintes de ce type de démence ont identifié des signes précurseurs et des symptômes typiques qui apparaissent à peu près dans l’ordre suivant.
La région de l’hippocampe dans les profondeurs du cortex temporal étant généralement la première touchée, il n’est pas étonnant de constater que les premiers symptômes consistent en des troubles de la mémoire. Ces derniers se manifestent initialement par des distractions mineures que l’on peut facilement confondre avec l’un des effets courant du vieillissement normal (oublier ses clés, un rendez-vous, le nom de quelqu’un, etc).
À ce stade léger (ou initial), les souvenirs plus anciens sont relativement préservés. Mais les oublis de choses récemment apprises deviennent fréquents et seront bientôt remarqués par les proches. Ceux-ci seront de plus en plus souvent questionnés, et parfois à plusieurs reprises, sur une date ou un événement à venir. On assiste aussi à l’utilisation plus fréquente de notes et d’aide-mémoire. Malgré cela, la victime de ces pertes de mémoire épisodique peut avoir tendance à les minimiser au début.
Parmi les autres symptômes cognitifs qui peuvent apparaître durant ce premier stade, on retrouve une durée d’attention plus courte, de la difficulté à se concentrer, des erreurs de jugement et des problèmes d’orientation spatiale ou temporelle. Les patients ont par exemple tendance à errer ou à se perdre dans des endroits qui leur sont pourtant familiers.
L’humeur et le comportement commencent aussi à être affectés, menant parfois à une exagération des traits de caractère de la personne (agressivité chez les colériques, passivité chez les placides, etc), à des sautes d’humeur, des états dépressifs ou à de la nervosité. La prise de conscience par moment de la détérioration de ses facultés peut devenir une source d’anxiété.
De légers problèmes de coordination motrice peuvent survenir, mais en général, à ce stade initial, la personne atteinte continue à mener une vie relativement indépendante avec un minimum d’aide.
On entre dans le stade modéré de la démence de type Alzheimer quand le jugement commence à être altéré au point où l’organisation des activités quotidiennes devient difficile. C’est le stade de la perte d’autonomie. La personne peut avoir par exemple de la difficulté à accomplir des tâches familières qu'elle a exécutées toute sa vie, comme préparer ses repas, faire sa toilette ou son budget.
Les facultés cognitives comme la mémoire continuent de se détériorer et la personne a de plus en plus de mal à apprendre et à retenir de nouvelles informations. Leurs souvenirs de jeunesse et d’âge moyen sont toutefois encore préservés.
Les plaques amyloïdes et la dégénérescence neurofibrillaire s’étendent progressivement aux régions impliquées dans le langage ainsi que dans la perception du corps parmi les objets qui l’entourent. D’où les difficultés dans les tâches quotidiennes, mais aussi dans la vie professionnelle et sociale qui souffrent énormément de la dégradation des facultés langagières : la moindre conversation devient laborieuse, la personne perd le fil de ses idées, ne trouve plus les mots pour s’exprimer, ne comprend pas bien ce qu’on lui explique. Elle délaisse alors au même moment la lecture et l’écriture.
D'un point de vue comportemental, les patients deviennent souvent irritables ou indifférents, ont des gestes stéréotypés, pleurent ou rient sans raison apparente, ont des gestes déplacés, etc. Il ne faut pas oublier que deux personnes évaluées au même stade peuvent présenter des atteintes différentes, les facultés cognitives pouvant être plus touchées que l’autonomie comportementale, ou l’inverse.
Au stade avancé, certains négligent souvent leur hygiène corporelle, peuvent oublier de manger, perdent le contrôle de leur vessie et de leurs intestins. À ce stade, une surveillance permanente par les proches ou l’hébergement dans un centre de soins devient nécessaire. Soutenir la personne atteinte pour lui assurer la plus grande qualité de vie possible devient alors l’objectif le plus important.
Avec le temps, la démence de type Alzheimer devient souvent une « démence mixte », c’est-à-dire qu’elle cohabite avec d’autres formes de démence, comme la démence vasculaire et la maladie à corps de Lewy. Le patient peut mourir d’une autre maladie à n’importe quel stade de l’évolution de sa démence. Cependant, dans le stade avancé de la démence de type Alzheimer, beaucoup de décès sont causés par une infection secondaire. Un cas fréquent étant la pneumonie engendrée par de la salive ou de la nourriture que le patient, qui a de la difficulté à avaler, laisse entrer dans ses voies respiratoires.
Celle-ci est divisée en 7 stades qui mesurent l'évolution de la démence de type Alzheimer, mais aussi d'autres démences, bien que les stades et les symptômes diffèrent selon chacune. Ces stades sont définis de manière qualitative, allant du fonctionnement normal ne présentant aucune atteinte (stade 1) au déclin cognitif très sévère (stade 7), en passant par des états intermédiaires où les facultés mentales sont qualifiées de très légèrement affaiblies, de légèrement affaiblies, de modérément affaiblies, etc.
La détermination des critères de ces stades comporte évidemment une part de subjectivité puisque de toute façon les atteintes n’évoluent pas selon une séquence stricte pour tous les patients.
Stade 1 : Stade de référence
Ne vit aucune difficulté dans le cadre de la vie quotidienne
Oublie les noms et l'emplacement de certains objets
Peut avoir de la difficulté à trouver ses mots
A de la difficulté à aller à de nouveaux endroits
A de la difficulté à faire face aux problèmes qui surviennent au travail
A de la difficulté à accomplir de tâches complexes (finances, emplettes, planification d'un repas avec invités)
A besoin d'aide pour choisir ses vêtements
A besoin d'être incité à se laver
A besoin d'aide pour s'habiller
A besoin d'aide pour, ou à peut-être peur de, prendre son bain
Est de moins en moins capable d'aller aux toilettes, ou est incontinent
Vocabulaire limité pour enfin ne répondre que par un seul mot
Perd la capacité de marcher et de s'asseoir
Devient incapable de sourire
Ainsi, les gens dont le diagnostic survient autour de 60 ou 65 ans déclinent généralement plus lentement et leur espérance de vie est alors plus de l’ordre de 12 ou 14 ans, certains allant même jusqu’à 20 ans. Par contre, ceux dont le diagnostic arrive à 80 ou 85 ans ont tendance à progresser plus rapidement. Leur âge avancé augmentant en plus leur probabilité de décès, ils ne survivent généralement qu’environ 5 à 8 ans.
Par ailleurs, la survie après le diagnostic a tendance à s'allonger, entre autres à cause de l'amélioration des méthodes diagnostiques, des meilleures conditions de prise en charge, et à mesure que les personnes sont plus disposées à se faire évaluer.
A + de 85 ans 4% souffrent de la maladie de Parkinson.
Cette situation pose donc la question de la qualité de vie des personnes âgées mais aussi de son coût pour la société aussi bien financier qu'en matière d'impact sur les accompagnements.
Après le sevrage (3 semaines), des souris transgéniques « Alzheimer » sont placées soit en milieu enrichi, soit en milieu standard pendant 5 mois (équivalant à 13 ans d'étude pour nous). Le milieu enrichi est constitué de grandes cages (6 à 8 souris par cage) avec des roues, des tunnels, des constructions ; plusieurs jouets en plastique sont mis et changés une fois par semaine 1. De plus, les souris de cette condition enrichie ont des sorties « culturelles » trois fois par semaine, et visitent librement toute sorte de dispositifs servant aux expériences, labyrinthes, tubes... Le milieu standard est constitué de cages habituelles mais avec les mêmes rations de nourriture et d'eau. Les tests sont variés (différents labyrinthes ou test de reconnaissance) et durent 6 semaines, soit un mois et demi.
L'effet du milieu enrichi est très bénéfique. Après 45 essais d'un apprentissage à trouver le but dans un labyrinthe (9 jours d'apprentissage à raison de 5 essais par jour), les souris transgéniques « Alzheimer » élevées en milieu enrichi sont aussi performantes que des souris saines (du milieu standard). Au premier essai d'apprentissage, toutes les souris mettent environ 45 secondes pour arriver au but. Au bout du 4e essai, les souris « Alzheimer » élevées en milieu enrichi atteignent le but en 20 secondes, soit aussi rapidement que les souris saines élevées en milieu standard. Le rappel 30 minutes après ce 4e essai donne un temps encore meilleur d'environ 15 secondes. L'environnement enrichi compense complètement le déficit cognitif lié à la maladie d' Alzheimer. A l'inverse, les souris « Alzheimer » élevées en milieu standard restent, après quatre jours d'entraînement, à plus de 30 secondes pour atteindre le but, c'est-à-dire qu'elles ne réussissent pas à apprendre vraiment. Enfin, les meilleures sont les souris saines élevées en milieu enrichi, atteignant le but en 10 secondes. La stimulation cérébrale est donc très efficace dans tous les cas.
Des analyses du cerveau sont réalisées sous microscope après l'expérience et révèlent des changements conséquents. Non seulement la surface de dépôt de plaques séniles (protéine A4-amyloïde) dans le cortex est moins importante quand les souris transgéniques « Alzheimer » ont été élevées en milieu enrichi mais ce dépôt est encore plus bas pour les souris « Alzheimer » qui ont été élevées en milieu enrichi et ont participé à tous les tests d'apprentissage.
Ces chercheurs trouvent les mêmes résultats sur la diminution des plaques séniles dans l'hippocampe (nécessaire à l'enregistrement en mémoire). Ainsi, leur calcul indique
une réduction de 57 % de plaques séniles dans le cortex et 69 % dans l'hippocampe chez les souris « Alzheimer», élevées en milieu enrichi et qui ont été testées. Autrement dit,
les apprentissages, notamment dans les labyrinthes, constituent un milieu super-enrichi (nouveauté) ou déclenchent, grâce à d'autres mécanismes (motivation, stress ?) une stimulation cérébrale supplémentaire.
Ce résultat me rappelle que lorsque, avec mon ami neurologue Hervé Allain, nous travaillions avec des étudiant(e)s dans des résidences pour personnes âgées, ces personnes étaient tout d'abord réticentes, ne voulant pas délaisser leur partie de cartes ou leurs feuilletons favoris. Mais après les expériences, c'étaient tout le contraire et ils demandaient aux étudiant(e)s quand ils reviendraient. Une conclusion essentielle de toutes ces études est que l'apprentissage en lui-même est une source très riche d'enrichissement cognitif.
Chaque fois, donc, que vous vous « cassez » la tête pour monter un meuble livré par petits bouts, que vous cherchez comment programmer une émission sur votre nouvel enregistreur DVD, ou que vous relisez des passages compliqués de ce livre... vous stimulez de façon bénéfique votre cerveau.
Comme dirait La Fontaine, « Nouveauté et difficulté font plus que routine et facilité !!! » (extraits du livre d'Alain Lieury "Doper son cerveau, réalité ou intox")
Ce test consiste à faire apprendre à un sujet les mots d'une liste puis de lui demander de se rappeler de ces mots.
- C'est à dire que dans le cas où il ne peut retrouver lui-même les mots de la liste, on lui donnera dans un premier temps un indice (=nom de la catégorie sémantique du mot oublié).
=> Exemple: le mot fleur sera donnée pour aider le patient à retrouver le mot marguerite.
Ce type d'aide s'appelle un rappel indicé.
- Si malgré cette première aide, l'individu ne retrouve pas le mot on va l'aider d'une autre manière en lui donnant d'autres mots que l'on appelle distracteurs (=mots appartenant à la même catégorie sémantique que le mot oublié).
=> Dans l'exemple, il s'agira ici de paquerette, coquelicot...
Ce type d'aide s'appelle un rappel en reconnaissance.
1 - Les plaques séniles (ou plaques amyloïdes) : Ces plaques, essentiellement localisées dans le néocortex et l'hippocampe, correspondent à l'accumulation extracellulaire d’un peptide dit « ?-amyloïde » (PbA) ou « peptide A?42 » (42, parce que constituée de 42 acides aminés). Cette protéine est en fait la forme clivée et anormale d'une glycoprotéine membranaire dite « protéine précurseur de la protéine ?-amyloïde » (ou APP). C'est une protéine pathogène (neurotoxique) et insoluble qui est aussi la forme la plus amyloïdogène du bêta-amyloïde.
2 - Les dégénérescences neurofibrillaires : Des lésions cellulaires semblent consécutives à une augmentation de la capacité de phosphorylation (estérification des fonctions alcool des acides aminés hydroxylés) de la protéine Tau (macromolécule essentielle à la stabilité de la tubuline (protéine constituant majoritairement l'assemblage des microtubules qui forment le cytosquelette des axones neuronaux)). Ces lésions semblent induites par l'accumulation anormale d'un dérivé de la protéine tau hyperphosphorylée, responsable de la formation de filaments appariés : quand la protéine tau est hyperphosphorylée, elle se détache des microtubules, et va se conformer en paire de filaments hélicoïdaux pathologiques, qui s'agrégent en amas de neurofibrilles causant une neurodégénérescence fibrillaire progressive. Les substances nécessaires au fonctionnement du neurone ne pouvant plus être acheminées jusqu'au corps cellulaire, le neurone finit par mourir.
Aux États-Unis les législateurs commencent à reconnaître, grâce aux images IRM, l'origine biologique de ces affections, le la possibilité d'en faire un diagnostic et de leur apporter un traitement...
Ce qui a été observé chez certains schizophrènes qui "entendent des voies" c'est que les voies de communication entre les lobes frontaux et temporaux sont anormales, en pparticulier du côté gauche.
On peut donc imaginer que, s'il y a un retard trop important entre la production interne de cette voix et sa perception, ou réverbération par les régions audidives, l'impression soit donnée au patient que la voix vient d'ailleurs. On pense aussi que le développement anormal de certaines connexions au début de la vie pourrait contribuer à l'autisme.
- Description inlassable des symptômes physiques ;
- Difficulté pour communiquer verbalement les émotions ;
- Discours répétitif ;
- Production fantasmatique et onirique pauvre ;
- Relations interpersonnelles marquées par une forte dépendance, généralement manifestée par une mise à distance.
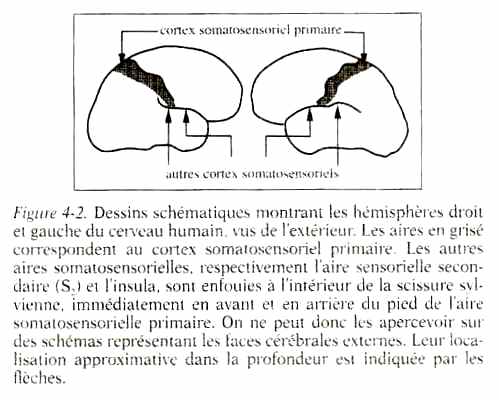
- Premièrement, il existe une région du cerveau humain, le cortex préfrontal ventro-médian, dont la lésion perturbe, de façon constante et très claire, les processus de raisonnement et de prise de décision, ainsi que ceux de l'expression et de la perception des émotions, surtout dans le domaine personnel et social. On pourrait aussi dire métaphoriquement que les processus neuraux sous-tendant la raison et les émotions se « recoupent » au niveau du cortex pré-frontal, ainsi qu'au niveau de l'amygdale.
- Deuxièmement, il existe une région dans le cerveau humain, constituée par un ensemble d'aires corticales somatosensorielles situées dans l'hémisphère droit, dont la lésion perturbe également les processus de raisonnement et de prise de décision, ainsi que ceux relatifs à l'expression et à la perception des émotions, et, en outre, interrompt la perception des messages sensoriels en provenance du corps.
- Troisièmement, il existe des régions situées dans le cortex préfrontal, outre le secteur ventro-médian, dont la lésion perturbe aussi les processus de raisonnement et de prise de décision, ainsi que ceux relatifs à l'expression et à la perception des émotions, mais selon des modalités différentes : soit le déficit est beaucoup plus large, impliquant les fonctions intellectuelles dans tous les domaines ;
soit il est beaucoup plus sélectif, touchant à la manipulation des mots, des nombres, des objets ou de l'espace, davantage qu'aux manifestations relevant du domaine personnel ou social. On peut voir une carte très simplifiée de ces importants recoupements dans la figure 4-4.
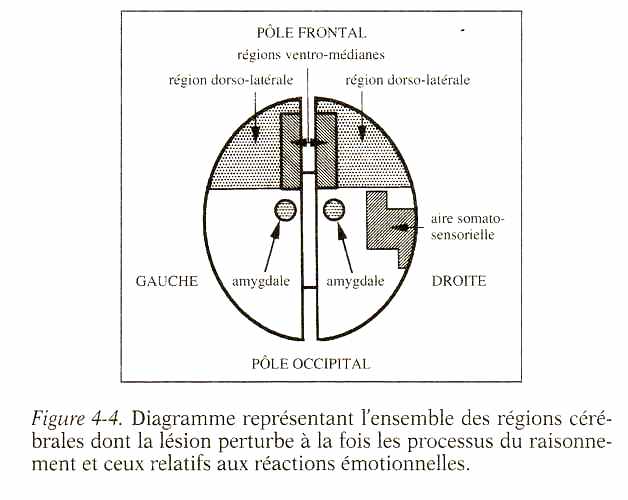
- Premièrement, ces systèmes sont certainement impliqués dans les processus de raisonnement, dans le sens large de ce terme. Plus précisément, ils sont impliqués dans les processus de planification de l'action et de décision.
- Deuxièmement, un sous-ensemble de ces systèmes neuraux est associé à la planification et à la prise de décision dans les domaines que l'on peut qualifier de « personnel et
social ». Il semble bien que ce sous-ensemble de systèmes soit lié à la mise en oeuvre d'actes que l'on qualifie généralement de raisonnables.
- Troisièmement, les systèmes neuraux que nous avons identifiés jouent un rôle important dans les réactions émotionnelles.
- Quatrièmement, ces systèmes neuraux sont nécessaires pour pouvoir maintenir présente à l'esprit, pendant une période de temps prolongée, l'image d'un objet pertinent, lorsque ce dernier est soustrait à la vue directe.
Les patients assistent, impuissants, à des mouvements très organisés de leur bras, par exemple, pour aller saisir un stylo ou un verre comme dans le syndrome de la main capricieuse. Il manque cette force de contrôle qui devrait empêcher l'exécution du mouvement. Assis à table devant un verre d'eau, ces patients s'en emparent pour le boire, même s'ils n'ont plus soif.
Ce n'est d'ailleurs pas seulement le bras qui échappe au contrôle conscient de l'individu, mais tout un système, car les régions motrices qui actionnent le bras ne peuvent pas saisir le verre sans la coopération active du système visuel.
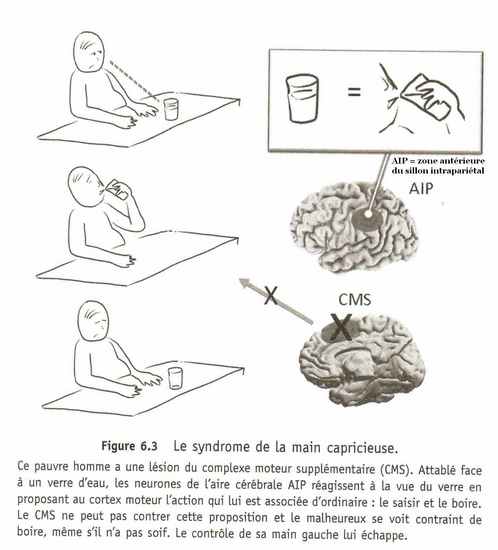
Si vous teniez devant vous une photo d'un voilier sur une mer démontée, vous n'auriez aucun mal à distinguer trois grandes régions dans l'image correspondant au bateau, au ciel et à la mer. Le bateau vous apparaîtrait lui-même composé d'une coque et d'une voile, etc. Tout cela semble très naturel. Pourtant, identifier les différents objets présents dans une scène est une véritable prouesse, un véritable casse-tête pour les ordinateurs. La coque et la voile du bateau sont-ils des objets en tant que tels, ou des parties de l'objet « bateau » ? Peut-on voir chaque vague comme un élément séparé ? Oui et non. La plupart des objets peuvent être vus comme des assemblages d'objets plus élémentaires ; inversement, il est souvent possible de regrouper des objets multiples au sein de groupes qui peuvent eux aussi être vus comme des objets. C'est le cas du bateau, mais aussi des bancs de poissons ou des vols d'oies sauvages. Il semble donc que l'on puisse débattre sans fin pour savoir si tel ou tel être ou chose est un objet ou bien un groupe d'objets. Pourtant, n'importe quel patient simultagnosique tranchera sans ambiguïté dans un sens ou dans l'autre. Pour le cerveau, il n'y a pas de débat. Les patients simultagnosiques parviennent plus facilement à voir plusieurs lettres en même temps si celles-ci font partie d'un même mot. Il peut même leur arriver de percevoir en même temps un mot et une image, si ceux-ci sont reliés par un lien sémantique, par exemple le mot « lion » écrit sur un dessin de lion. La simultagnosie nous renseigne donc sur ce qu'est, pour le cerveau, un objet, elle nous permet de comprendre ce qui est un et ce qui est deux.
TROUBLES DE LA VISION
L'imagerie fonctionnelle cérébrale peut nous montrer cette région en pleine action chez des gens bien portants. Au cours d'une expérience utilisant la TEP, et plusieurs fois répliquée depuis, Semir Zeki a présenté d'une part des Images abstraites vivement colorées (dans le style du peintre Mondrian), et d'autre part les mêmes images en noir et blanc. Il a ensuite cherché quelles régions cérébrales étaient plus fortement activées par les images colorées que par les images en nuances de gris. Une petite région, à la face inférieure des lobes occipitaux, baptisée V4, remplissait cette condition. Il est bien probable que cette région, activée par les couleurs dans un cerveau normal, n'est autre que celle dont la destruction accidentelle rend achromatopsique.
Cette pathologie est généralement due à une lésion de la macula correspondant à une légère fossette située au centre de la rétine et responsable de l'acuité visuelle. C?est à cet endroit que l?acuité visuelle est maximale. La macula correspond à une partie de la rétine ne contenant aucun vaisseau. Sa vascularisation est assurée en profondeur par la choroïde, qui est la membrane sous-jacente à la rétine, et en périphérie par les vaisseaux de la rétine elle-même.
Le patient voit les lignes droites ondulées, incurvées ou brisées et les lettres déformées. Si, d'autre part il signale une diminution de la taille des images, une tache grise dans le champ visuel, une baisse de l'acuité visuelle et une diminution de la taille des images, il existe alors une forte présomption d'un syndrome (ensemble de symptômes) maculaire. Celui-ci est quelquefois secondaire à la présence d'?dème (collection liquidienne)d'hémorragies ou d'une diminution de la substance entrant dans la composition de la macula (dégénérescence).
Les différentes régions de la mosaïque du système visuel, V4, V5 et les autres, sont-elles distribuées n'importe où dans le cerveau? Prenons un peu de recul. Ce que nous attendons d'un système de perception comme le système visuel ou le système auditif, c'est de nous informer sur ce qui se trouve autour de nous, sur la scène dans laquelle nous sommes plongés. Or connaître ce qui nous entoure, cela revient d'une part à savoir quelles sont les choses qui nous environnent (des chaussures, le chien, le soleil, le commissariat de police...), et d'autre part la position, l'orientation, le mouvement de ces choses par rapport à nous. Ungerleider et Mishkin ont suggéré, il y a près de vingt ans, que ces deux aspects de la vision (qu'on peut résumer en deux mots : QUOI et OÙ) ressortiraient schématiquement de deux secteurs distincts du cortex visuel (Figure 13). Le QUOI relèverait de la partie la plus basse du cortex visuel, placée sur le dessous du cerveau dans les lobes occipitaux et temporaux. La région des couleurs, V4, dont nous venons de parler, fait partie de ce premier ensemble. En revanche, le OÙ, c'est-à-dire les aspects de la vision qui permettent de localiser les objets dans l'espace, de diriger les yeux ou la main vers eux, relèverait de la partie la plus haute du cortex visuel, dans les lobes occipitaux et pariétaux. La région du mouvement, V5, appartient plutôt à ce second ensemble.
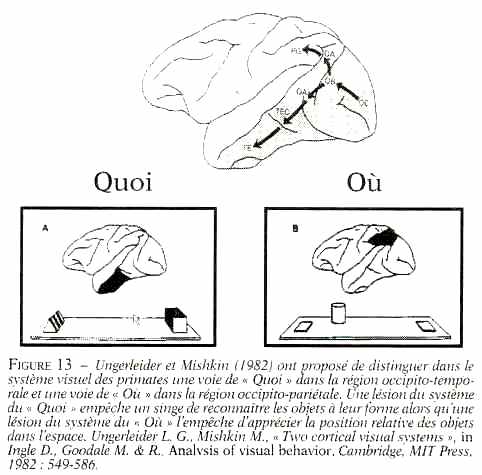
Le laboratoire de McGaugh s'est longtemps préoccupé du rôle des hormones périphériques comme l'adrénaline dans la consolidation des processus de mémoire. Ses études montrent que si des rats reçoivent une injection d'adrénaline juste après un apprentissage, ils ont un souvenir accru de leur activité à ce moment-là. Cela suggère que si l'adrénaline est libérée naturellement (des glandes surrénales) dans une situation donnée, le souvenir de cette situation reviendra plus facilement en mémoire. Comme l'activation émotionnelle induit généralement la libération d'adrénaline, on peut s'attendre à ce que les souvenirs explicites, conscients, des situations soient plus forts lorsque ceux-ci sont riches en émotions. Et on peut aussi prévoir que le blocage des effets de l'adrénaline neutralise les effets stimulants sur la mémoire de l'activation émotionnelle.
McGaugh et Larry Cahill ont testé ces hypothèses. Ils ont demandé à des gens de lire l'histoire d'un garçon faisant du vélo. Pour certaines personnes, le garçon faisait un tour puis rentrait et partait avec sa mère en voiture à l'hôpital chercher son père, un médecin. Pour d'autres personnes, l'enfant faisait un tour de vélo, était renversé par une voiture et emmené à l'hôpital où travaillait son père, un médecin. Les deux histoires étaient très proches par les mots utilisés et différaient uniquement par la manipulation des émotions induites. Après avoir lu les histoires et avant d'être testés sur leurs souvenirs, la moitié des sujets dans chaque groupe recevait soit un placebo, soit une substance bloquant les effets de l'adrénaline. Parmi les sujets ayant reçu le placebo, ceux qui avaient lu l'histoire émotionnelle se rappelaient plus de détails que ceux qui avaient lu l'histoire anodine. Mais chez ceux qui avaient reçu la substance bloquante, il n'y avait aucune différence de souvenirs entre les deux histoires, les deux groupes se comportant comme celui avec placebo et l'histoire non émotionnelle. Le blocage de l'adrénaline empêchait bien les effets stimulants de l'activation émotionnelle sur la mémoire.
McGaugh a suggéré quelques applications pratiques de ces résultats fascinants. Lors des conflits, les secouristes et les soldats sont souvent traumatisés par le souvenir des scènes horribles auxquelles ils ont assisté. Il serait peut-être possible, immédiatement après de telles expériences, de bloquer chez ces gens les effets de l'adrénaline et de leur épargner quelques angoisses plus tard.
Dans des situations de stress, le cerveau produit davantage d'enképhaline, ce qui masque parfois des lésions en cours. Par exemple, certains athlètes ne sentent pas la douleur d'une blessure pendant l'effort et ne prennent conscience de leur blessure qu'une fois la compétition terminée, quand leur stress est redescendu. Les scientifiques ont aussi trouvé que certains médicaments pouvaient bloquer la libération d'enképhaline, un blocage qui a pour conséquence d'augmenter la douleur.
Les enképhalines sont des endorphines fabriquées par le corps. Ce sont de petites protéines produites par des interneurones spécialisés. Ces derniers sont en communication avec les neurones de la douleur. Les enképhalines se fixent sur des récepteurs opioïdes présents à la surface de la membrane des neurones de la douleur. Elles inhibent les messages de la douleur (messages nociceptifs), vers le cerveau.
L'enképhaline est un neurotransmetteur qui n'est pas seulement spécifique à la douleur. Il a comme fonction principale de moduler le message nerveux. Il réduit la douleur, mais aussi la quantité de dopamine produite, c’est-à-dire qu'il intervient dans la variation de l'intensité du plaisir (sensation de plaisir venant donc des neurones à dopamine).
Les enképhalines ont une très courte durée de vie : quelques minutes en général. Elles sont normalement rapidement dégradées par des enzymes (hNEP, hAP-N).
La venue du sommeil est donc liée à l'augmentation du neurotransmetteur Gaba. C'est pour cette raison que les somnifères sont des médicaments qui facilitent la transmission des Gaba, en favorisant certains récepteurs des neurones à neurotransmetteur Gaba.
Rôle du glutamate dans la mémoire.
On trouve le corps cellulaire de tels neurones dans une zone bien précise du système nerveux central : l'hypothalamus et plus précisément dans le noyau tubéromamillaire de l'hypothalamus postérieur. Ce noyau, situé à la périphérie des corps mamillaires, est formé de cellules de grandes tailles qui se projettent de façon diffuse sur le cortex cérébral et en particulier sur les régions ventrales (hypothalamus, le prosencéphale basal, amygdale) qui sont fortement innervées.
Les axones de ces neurones histaminergiques, largement répartis dans tout le cerveau, y libèrent de l'histamine, qui va se fixer sur les récepteurs H1 et H2 de neurones, tous postsynaptiques et des récepteurs H3 présynaptiques. Ces derniers sont des autorécepteurs présynaptiques inhibiteurs. Les récepteurs H3 peuvent être aussi postsynaptiques et jouer le rôle de régulateurs de la libération des autres neurotransmetteurs. Ils sont largement distribués dans le striatum, le noyau accumbens et le cortex cérébral.
Complément : Le monoxyde d'azote, oxyde azotique, oxyde nitrique ou NO est un composé chimique formé d'un atome d'oxygène et d'un atome d'azote. C'est un gaz dans les conditions normales de pression et de température. C'est un important neurotransmetteur chez les mammifères et le seul neurotransmetteur gazeux connu.... À la différence de la majorité des autres neurotransmetteurs, dont l'action dans la fente synaptique a pour cible unique le neurone post-synaptique, la petite molécule de NO diffuse largement et peut atteindre plusieurs neurones environnants, y compris des neurones non inter connectés par des synapses. On pense que ce processus est impliqué dans la mémorisation en assurant la mémorisation à long terme. Les endocannabinoïdes, neurotransmetteurs lipidiques, ont des propriétés de diffusion similaires.
L'événement déclencheur initial, soit ici l'arrivée à maturation du foetus, est donc à l'origine de toute une série de circuits spécifiques, engendrant des comportements spécifiques, ceux-ci étant identiques de génération en génération. En ce sens les circuits peuvent être considérés comme des « mémoires ancestrales ». Bien évidemment, ces « mémoires » ont permis une meilleure adaptation de l'animal.
L'activation d'un circuit est toujours associée à une valorisation. Ainsi, la mère est heureuse d'allaiter ses petits.
La femelle campagnol qui produit du lait éprouve un plaisir physique, et donc une valorisation, à allaiter. Par ailleurs, grâce à l'apprentissage associatif, la production de lait et l'allaitement sont étroitement associés à la perception visuelle, sonore ou olfactive de petits. Donc, pendant des générations, il y a eu association entre la production de lait, le plaisir d'allaiter, une valorisation positive, la vision des petits, le fait de les lécher, etc. : ainsi, l'animal était toujours heureux en présence de ses petits.
Avec l'évolution, il est probable que la sécrétion d'ocytocine fut non seulement associée directement aux comportements maternels précédents, mais aussi à un circuit de valorisation positive, et cela de manière innée. Tous ces circuits furent transmis de génération en génération, ce qui expliquerait, après l'accouchement, l'intérêt immédiat de la femelle pour ses petits, c'est-à-dire l'instinct maternel.
Des expériences sur les rats campagnols ont montré l'importance incroyable des neurotransmetteurs dans leur comportement. On a injecté de l'ocytocine dans le cerveau d'une femelle campagnol vierge : elle est devenue immédiatement très « maternelle » avec des petits (qui ne sont évidemment pas les siens) ; elle les lèche, les protège... Inversement, si l'on supprime la circulation de l'ocytocine dans le cerveau d'une rate venant d'avoir des petits, son instinct maternel disparaît immédiatement : elle se désintéresse totalement de ses enfants... On constate la même chose pour une brebis !
On peut donc postuler que l'injection d'ocytocine active plusieurs circuits liés à l'instinct maternel, et permet l'augmentation des valorisations liées à ces circuits : par exemple, le circuit de « l'action de lécher », le circuit de « telle odeur », etc.
La sérotonine – ou hydroxytryptomine (5-HT) – a été identifiée pour la première fois il y a plus de 50 ans et, depuis, les scientifiques lui découvrent sans arrêt un nouvel intérêt, une nouvelle fonction dans l’organisme.
Sérotonine et autres molécules impliquées dans la dépression
Sérotonine et autres molécules impliquées dans la dépression - suite
animation des principaux noyaux émetteurs de neurotransmetteurs
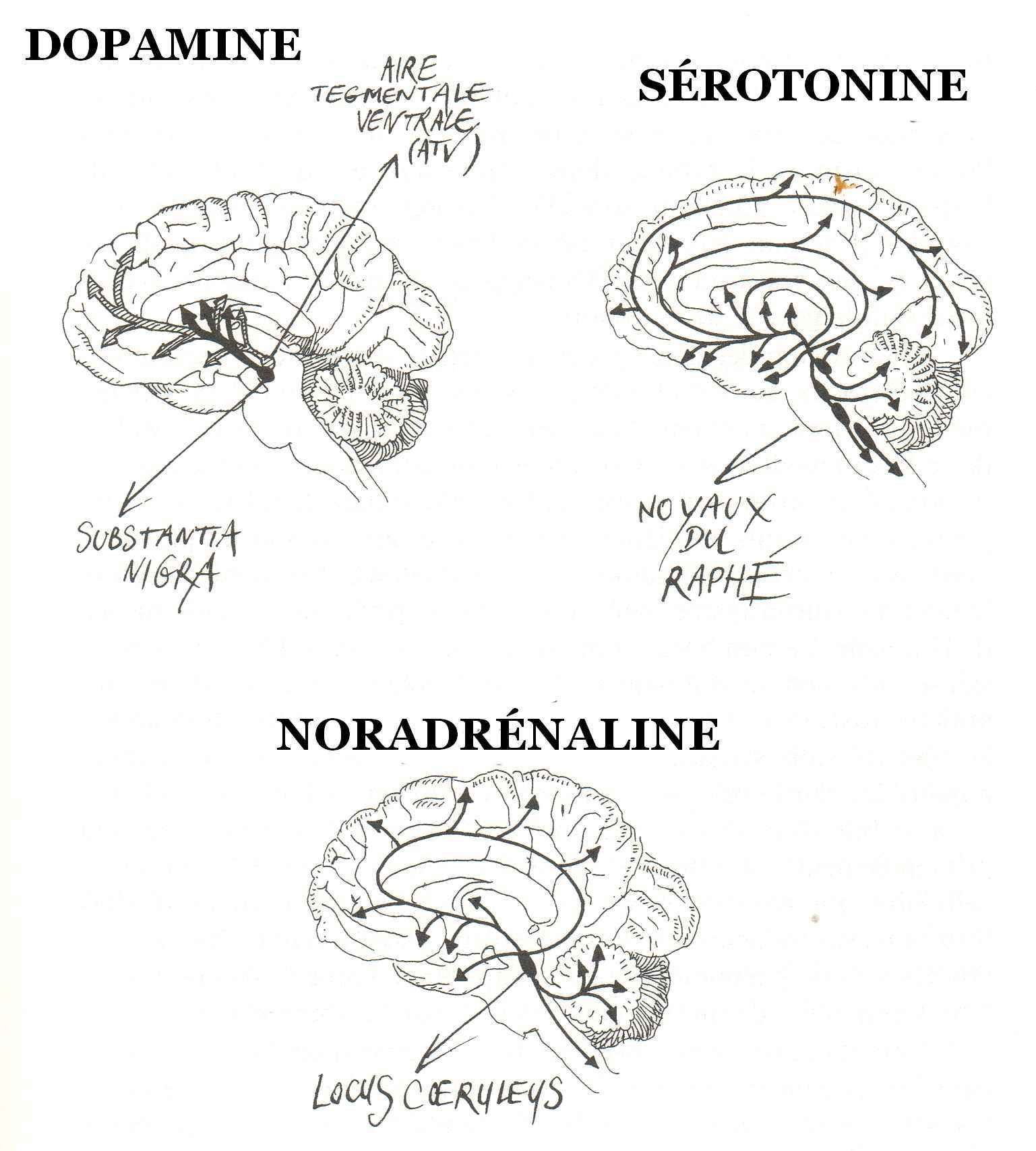
1º) Prendre conscience et identifier les situations qui ACTIVENT votre habitude de vous mordre les ongles. Par exemple : Caresser, toucher, palper, frotter ou gratter les ongles, les bords des ongles et les cuticules avant de porter vos doigts à la bouche. etc. (signalez tout ce que vous pouvez vous rappeler)
2º) Prendre conscience et identifier les situations qui PROVOQUENT ou STIMULENT votre habitude de vous ronger les ongles. Par exemple : En étudiant, moments d'examens, lire des livres, regardant la TV, face à l'ordinateur, discussions, état de grande nervosité, etc. (signalez tout ce que vous pouvez vous rappeler).
3º) Prendre conscience et identifier les situations où vous aviez ÉVITÉ votre habitude de vous mordre les ongles. Par exemple : En pratiquant des activités sportives, pendant des conversations avec des amis, en dansant, quand vous êtes dans des endroits publics, écouter et admettre les reproches de parents ou d'amis quand vous vous mordez les ongles, quand vous mastiquez quelque chose, etc. (signalez tout ce que vous pouvez vous rappeler).
4º) Effectuer des autocritiques constantes de la situation de vos ongles.
Par exemple : J'ai les ongles très laids, j'ai les doigts tordus, c'est une souffrance et une honte en voyant mes doigts et mes ongles, j’ai les ongles horribles, ect. (Signalez tout ce que vous pouvez vous rappeler).
Prendre conscience signifie percevoir des actions ou des situations que nous faisons consciemment. Dit d'une autre manière, une personne consciente se rend compte de ce qu'elle fait, tandis que la personne inconsciente agit avec des gestes ou des actes qui ne sont pas pensés consciemment. Si une personne fait un geste pour porter son doigt à sa bouche et mordre son ongle, ce n'est pas un acte conscient si elle ne le fait pas volontairement, parce qu’elle ne se rend pas compte tout ce qui affecte ses actions. En un mot, elle ne sait pas ce qu’il lui arrive.
La carte de saillance est le résultat de calculs automatiques menés au sein des aires sensorielles. Dans le cas de la vision, Les neurones de régions comme VI., V4 ou V5 analysent rapidement l'image - en moins de deux dixièmes de seconde - pour en extraire les contours des différents objets, leur vitesse, leur couleur et toutes sortes d'informations utiles. Ce processus d'analyse transforme l'image rétinienne, qui n'est, il faut le rappeler, pas grand-chose d'autre qu'une série de valeurs de luminance organisées selon deux dimensions - du rouge clair ici, du bleu foncé là - en un ensemble de régions homogènes vaguement reconnues comme « faisant penser à une tasse de café » ou « faisant penser à un visage ». La carte de saillance est l'un des résultats de ce processus de digestion de l'image. Elle favorise les éléments de l'image qui se démarquent nettement de leur environnement immédiat.
Les mécanismes neuronaux déterminant la saillance commencent à être suffisamment bien compris pour être reproduits sur ordinateur, au point qu'il existe même des programmes capables d'identifier les zones les plus saillantes d'une image. Si vous y réfléchissez, c'est un peu inquiétant ; cela signifie, en théorie, qu'une machine peut prédire quels éléments de l'image vont capturer votre attention en premier... et le libre arbitre dans tout ça ? Soyez tout de suite rassuré : ces programmes sont encore très approximatifs ; mais les progrès faits dans ce domaine illustrent tout de même à quel point notre attention est sensible, et dans une certaine mesure « sous le contrôle » de notre environnement. L'attention obéit à des lois, et celui qui les connaît peut dans une certaine mesure la contrôler.