
accueil

sommaire Science
autres textes 2005
|
accueil |
sommaire Science |
autres textes 2005 |
Certes, la sélection naturelle sélectionne les
espèces, mais comment se fait-il que de nouvelles espèces
apparaissent, dérivant des précédentes, et qui
pourront ensuite être sélectionnées si elles
s'avèrent aptes à survivre ?
Dans la conception habituelle du "programme génétique",
il est souvent invoqué des "erreurs accidentelles" dans la
recopie du code génétique : par une "erreur" dans la
recopie de ses chromosomes, un être inhabituel va naître,
et, s'il s'avère viable, il sera à l'origine d'une
nouvelle espèce (  note 1).
note 1).
Mon avis est qu'il ne faut pas concevoir le développement d'un
être vivant comme le déroulement d'un "programme"
génétique, analogue à un programme informatique,
mais comme une dynamique qui se déforme elle-même au fur et à mesure qu'elle grandit
et qui, parce que ses données de départ sont toujours
identiques, toujours se déforme de la même façon,
étape après étape, et produit, par
conséquent, toujours un même organisme à
l'arrivée.
Je vais essayer d'abord d'expliquer cette notion de dynamique qui se
déforme sur elle-même au fur et à mesure qu'elle
grandit, et l'on verra plus tard comment cela implique, très
simplement et même nécessairement du fait de
légères différences qui se font jour
inévitablement lors de la croissance de la dynamique, des
modifications de comportement qui peuvent s'amplifient jusqu'à
donner naissance à une nouvelle espèce.
L'idée de programme génétique a l'avantage
d'expliquer pourquoi tous les êtres d'une même
espèce se ressemblent : on déroule le même
programme, c'est-à-dire la même séquence
d'instructions, et, très normalement, à l'arrivée
on obtient toujours la même chose.
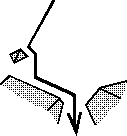
| Dans le cas d'une simple dynamique, on peut retrouver cette notion de répétitivité. Par exemple, si l'on considère une rivière qui dévale la pente d'une montagne (croquis ci-contre), toujours elle sera déviée au même endroit par les mêmes rochers ou par les mêmes changements dans l'angle de la pente de la montagne, et toujours elle arrivera donc en bas au même endroit. Même si l'absence de pluie pendant une longue période vient à assécher la rivière, lorsque la pluie reviendra elle retrouvera "toute seule" son chemin, chemin qui sera toujours le même chemin, sous réserve du moins que son débit soit à peu près équivalent. | |
 |
Dans cet exemple, on peut toutefois penser que la disposition de la
montagne est l'équivalent d'un programme qui se conserve et qui
informe la rivière à chaque passage sur ce qu'elle doit
faire. La dynamique de la rivière est effectivement
déformée ici par quelque chose qui lui est
extérieur, alors que l'on était parti pour évoquer
le cas d'une dynamique qui se déforme elle-même.
Pour donner un exemple d'une telle dynamique "auto-déformante",
nous allons rester dans le cas d'un fluide qui descend une montagne,
mais cette fois nous allons supposer qu'il s'agit de la lave d'un
volcan qui s'écoule lentement le long de sa pente. Pour que la
forme de la montagne n'influe pas sur le mouvement de la lave, on va
supposer que la montagne a la forme d'un cône parfaitement
régulier.
Donc, la lave s'écoule régulièrement. Selon sa
température en haut du volcan, sa fluidité sera plus ou
moins grande, et sa vitesse sera en conséquence plus ou moins
grande. À température égale, même
fluidité et même vitesse : cela est strictement
déterministe (croquis A).
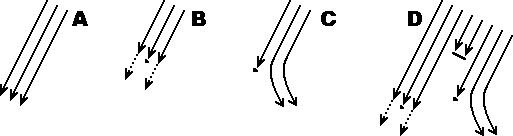
Supposons maintenant que la lave contienne une impureté interne,
c'est-à-dire, par exemple, un fragment de roche qui sera
tombé dedans. À son endroit, la lave va se refroidir plus
vite qu'alentour. Si l'on suppose aussi que la lave était juste
à la température lui donnant suffisamment de
fluidité pour s'écouler le long de la pente, à
l'endroit de l'impureté, la lave va s'arrêter. Si
l'impureté est située vers le centre de la coulée
de lave, celle-ci va "toute seule" se diviser en deux, les deux
branches se séparant à l'endroit de la lave qui est trop
refroidie par l'impureté et qui refuse, pour cette raison, de
descendre plus bas. Par ailleurs, puisqu'elle est divisée en
deux, la lave a perdu de sa compacité, ce qui fait que sa
température va baisser plus rapidement que si elle était
restée compacte. En conséquence, ces deux
demi-coulées en lesquelles elle s'est divisée vont
descendre moins bas dans la pente que si elle était
restée en une seule coulée compacte, car le moment
arrivera plus vite où leur température ne leur procurera
plus une fluidité suffisante pour continuer (croquis B).
On peut envisager un autre cas de figure (croquis C)où la
même impureté, au lieu d'être située vers le
milieu de la coulée de lave, se situerait près d'un bord.
Alors, au lieu de se diviser de part et d'autre de ce "point froid",
elle serait seulement détournée, déviée par
lui. Alors aussi, elle resterait compacte et descendrait la pente
pratiquement aussi bas que si elle avait continué tout droit.
Ainsi, dans cet exemple, la pente de la montagne ne donne aucune
information à la lave sur ce qu'elle doit faire, sur le chemin
qu'elle doit prendre et sur quelle longueur elle peut poursuivre :
toute la dynamique de la lave est seulement conditionnée par la
présence ou non d'une impureté en son sein et sur
l'endroit précis où se trouve cette impureté.
Cette seule présence suffit pour déterminer si la lave
ira loin ou si elle s'arrêtera plus rapidement, pour
déterminer si elle continuera en une seule branche ou si elle se
divisera en plusieurs, pour déterminer si elle arrivera à
tel endroit ou bien à tel autre.
Bien entendu, tout cela est également parfaitement
déterministe, et toujours la lave arrivera au même endroit
et de la même façon si une même impureté
existe en elle au même endroit.
On peut aussi raffiner un peu cet exemple.
On peut supposer, par exemple, que la lave contienne d'autres
impuretés plus petites (croquis D). Dans ce cas, ces
impuretés pourraient n'avoir aucune influence sur la lave
compacte, mais elles pourraient avoir une influence sur les branches de
la lave lorsqu'elle est divisée et donc plus rapidement
refroidie. Ces plus petites impuretés pourraient alors diviser
en branchioles les deux branches de la lave, et, selon encore la
position des petites impuretés dans la lave, ce sont
différentes figures qui en résulteront pour
l'écoulement de la lave.
Ce que l'on veut suggérer, c'est que le développement
d'un être vivant serait analogue à une coulée de
lave déformée par des impuretés internes. Dans le
cas d'un être vivant, c'est son développement même,
c'est-à-dire sa croissance, l'augmentation du nombre de ses
cellules, qui donnerait le rôle moteur que procure la pente de la
montagne dans le cas de la lave.
Dans l'hypothèse que je propose ( aller à ce texte),
un être vivant ne serait fondamentalement rien d'autre qu'un
courant électromagnétique qui fonctionne en boucle
fermée, et cela vaudrait jusqu'au coeur de chaque cellule dont
le chromosome ne serait rien de plus qu'un assemblage de bobines
électromagnétiques supraconductrices se repoussant au
moment de la division cellulaire (nota : j'insiste sur le
caractère supraconducteur que je prête à ce
courant, car c'est cette particularité qui le rendrait
usuellement indétectable, sauf à l'occasion de ses
perturbations).
Dans cette hypothèse, au moment des premières divisions
de la cellule qui donnera naissance à un être vivant, la
dynamique qui correspond à la lave dans l'exemple donné
plus haut serait donc tout simplement le courant
électromagnétique qui parcourt ses chromosomes, et les
impuretés, qui font que l'embryon va se développer de
telle façon plutôt que de telle autre, ne seraient autres
que les gènes répartis dans les chromosomes et dont le
courant électromagnétique propre détourne le
courant électromagnétique d'ensemble vers telle ou telle
direction. Je rappelle seulement que ( aller à ce texte),
dans ce rôle, le chromosome dit "poubelle" aurait un rôle
aussi essentiel que les parties dites "codantes" car, le chromosome
étant une construction dans les trois dimensions de l'espace,
selon la façon dont le chromosome "poubelle" sera
organisé dans l'espace, tel ou tel site "codant" sera plus ou
moins favorisé, c'est-à-dire plus ou moins placé
au coeur de la dynamique électromagnétique ou plus ou
moins marginalisé et donc rendu inopérant.
Pendant toute la vie de l'être vivant, ses cellules vont
d'ailleurs continuer à fonctionner sur ce principe, et des
conformations différentes dans l'espace du chromosome "poubelle"
expliqueront (selon mon hypothèse, du moins) pourquoi, selon les
organes, les cellules feront des choses différentes les unes des
autres, bien qu'elles aient toutes le même "code
génétique", c'est-à-dire les mêmes
gènes "codant".
À l'échelle de l'embryon en train de se former, ce qui se
passe au niveau de chaque cellule sera cependant bientôt en
concurrence (ou en osmose amplificatrice) avec le courant
électromagnétique qui circule dans l'ensemble de
l'embryon.
Mon hypothèse est que c'est ce courant d'ensemble qui
modèle progressivement l'embryon, le guide dans sa croissance,
et que, de la même façon que des impuretés
déformaient le courant de la lave en fusion donnée en
exemple, ce seront les
hétérogénéités qui se produiront
dans le courant électromagnétique d'ensemble de l'embryon
qui le déformeront dans tel ou tel sens, qui permettront
à tel ou tel organe de grandir ou à tel autre de cesser
sa croissance, ou à tel autre encore de changer de position et
de s'enfouir au coeur de l'embryon alors qu'il avait commencé
par apparaître à sa surface.
Comment se forment ces hétérogénéités ? Il est connu que ( lien vers un exemple de complexité en évolution),
du moins pour ce qui concerne l'organisation des axes
avant/arrière et dessus/dessous, c'est par la formation d'un
gradient de certaines substances que des différences se forment
entre les divers endroits de l'embryon.
Peut-être de subtiles différences de température ou
de proximité de la source d'approvisionnement en énergie
expliquent-elles l'origine de ces gradients.
Mais, plutôt que le hasard des circonstances qui font que tel ou
tel endroit de la cellule est mieux approvisionnée en
énergie, probablement ces gradients sont-ils l'effet de
l'hétérogénéités du courant
électromagnétique propre aux chromosomes. On sait, en
effet, que les gènes dit "homéotiques"qui correspondent
à l'ordre d'émergence et de disposition des
différents segments du corps (tête, tronc, arrière)
sont répartis dans le même ordre sur les chromosomes. Sous
cet aspect, les chromosomes apparaissent donc comme un petit être
vivant en réduction, et, du moins au départ, on peut donc
considérer que les
hétérogénéités entre ces divers
tronçons se contentent de s'amplifier en même temps que
grandit l'embryon, juste comme un courant qui grossit doucement et dont
les circonvolutions de détail deviennent progressivement plus
nettes, plus accusées.
Ensuite, on peut supposer que les chromosomes qui sont situés
dans des ambiances différentes voient leurs chromosomes
"poubelle" déformés de différentes façons
selon, par exemple, qu'ils sont dans des cellules situées dans
la zone "tête" ou dans des cellules situées dans la zone
"queue". Alors, selon les gènes qui seront ainsi mis en position
externe et ceux qui seront enfouis au fond du noyau par les
"chromosomes poubelle", des protéines différentes seront
formées. De la sorte, des
hétérogénéités de plus en plus
locales seront créées, modifiant de façon de plus
en plus fine le flux électromagnétique et modelant ainsi
de façon de plus en plus fine les organes du futur foetus.
À un moment donné, on peut penser qu'un basculement se
fait : c'est le moment où le coeur et les organes en
général sont suffisamment mûrs pour
générer un cycle énergétique autonome
basé sur l'alimentation depuis le cordon ombilical et sur
l'évacuation des déchets qui en résultent. Pour
modeler la suite du développement du foetus, le flux
énergétique des organes (notamment, leur
approvisionnement par le moyen du sang qui circule dans les veines et
les artères, mais aussi leur interdépendance au
système nerveux qui s'installe) prend alors le dessus sur son
flux électromagnétique.
Un point mérite d'être souligné cependant. La
règle est en effet que tout niveau de complexité se
construit sur les niveaux précédents mais qu'il ne les
fait pas disparaître pour autant ( lien vers un développement de cette idée)
: ainsi, la complexité de la vie organique n'a pas
détruit le niveau de complexité des atomes et des
molécules, niveau apparu plus tôt dans l'évolution
de l'univers, mais elle s'est construite dessus et exige que les
molécules continuent à se comporter en molécules
pour perdurer elle-même en tant que vie organique. De la
même façon, dans l'adulte vivant, l'organisation en
organes n'a probablement pas détruit le fonctionnement en flux
purement électromagnétique organisé dans l'embryon
et doit seulement tenir comme "au-dessus" de lui, en tant que simple
strate supplémentaire de complexité. Mon hypothèse
est que c'est ce niveau "antérieur" de fonctionnement du corps
vivant, celui qui était son fonctionnement
caractéristique à son époque embryonnaire, qui
correspondrait aux flux que l'acuponcture chinoise révèle
et influe. Cela expliquerait pourquoi, par le biais des
"méridiens" repérés par l'acuponcture, certains
organes apparemment éloignés fonctionnellement sont en
fait en communication directe et s'équilibrent mutuellement :
ils étaient proches dans l'embryon où ils
équilibraient déjà ensemble leur flux
électromagnétique dans un cycle particulier, de telle
sorte que ce fonctionnement continue tout simplement dans l'adulte.
Revenons à l'embryon.
Que ce soit le flux électromagnétique initial des
chromosomes ou que ce soit le flux de la croissance cellulaire
occasionnée par la division rapide des cellules, l'embryon est
donc, selon mon hypothèse, essentiellement une dynamique. Quand
elle fonctionne ou quand elle fait sa croissance, cette dynamique est
influencée par les "impuretés" que sont les points durs
ou au contraire les courants favorables qui la parsèment du fait
de son histoire précédente, c'est-à-dire du fait
des zones de blocage ou de fort courant qui sont nés des
hétérogénéités de sa croissance
précédente.
Comme il en allait pour la lave à la limite de s'arrêter
et dont une petite impureté suffisait à modifier
radicalement le cours, une petite
hétérogénéité, située
plutôt ici plutôt que là, suffit à modifier
le développement du flux de croissance de l'embryon.
Mais plus que la position des
hétérogénéités, peut-être la
force du courant de croissance est-elle essentielle, ou tout du moins
complémentaire ? Dans le cas de la lave, pour qu'une petite
impureté ait de l'importance sur son écoulement, il faut
que la température de la lave soit "limite". C'est pareil pour
le courant de la rivière qui dévale la montagne : si la
force du courant est grande, elle franchira sans sourciller l'obstacle
d'un barrage de rochers, tandis que, si le courant est faible, elle
devra changer son trajet pour les contourner.
Ainsi, dans l'embryon, il faudrait tenir compte de la force de son
courant électromagnétique ou de son courant de croissance.
Cette force est probablement liée à la quantité d'énergie que peut recevoir l'embryon.
Dans le cas d'une espèce qui se reproduit au moyen d'un oeuf,
cette énergie est d'abord donnée par la qualité de
la nourriture qui a été incluse dans l'oeuf avant la
formation de sa coquille, puis par la quantité de chaleur que
recevra ensuite l'oeuf au cours de son développement. Et l'on
sait que, dans le cas des crocodiles et des tortues, la
différence de température apportée à l'oeuf
est la cause qui suffit à faire, selon les cas, des mâles
ou des femelles.
Dans le cas d'une espèce où le foetus se développe
dans le corps de la mère, cette énergie dépend
tout le temps de la nourriture que la mère peut lui apporter, et
donc a priori de la quantité et de la qualité de la
nourriture que reçoit la mère elle-même.
Ainsi, on peut supposer, par exemple, que dans un environnement
où la nourriture se raréfie durablement, le flux de
croissance de l'embryon soit ralenti pour se caler sur le rythme plus
lent avec lequel l'énergie lui parvient. Dans le cas d'une
rivière, on sait que les modifications de son débit ne
modifient pas systématiquement son cours et que cela arrive
seulement à certains débits qui sont critiques
vis-à-vis des obstacles qu'elle rencontre. De la même
façon, dans le cas d'un embryon, on doit penser que les
variations de l'énergie qu'il reçoit ne modifient pas
systématiquement la forme de son développement, et donc
la forme qui résultera de son développement, mais que
cela peut se produire dans certaines situations critiques, ou pour le
développement de certains organes critiques.
Peut-être le ralentissement du "débit de croissance"
va-t-il assécher la croissance de tel organe au détriment
de tel autre, ou bien au contraire va-t-il laisser plus de temps
à tel ou tel organe pour se complexifier. Ou bien un blocage se
produira localement, qui bouchera complètement une direction de
développement et qui forcera le développement à se
détourner vers une autre direction ou à se diviser en
plusieurs branches. Qui sait ce que donneront ces modifications dans la
croissance de l'embryon ? Peut-être un être monstrueux qui
ne sera pas viable, mais peut-être, de temps en temps, quelque
chose de parfaitement viable et que le hasard aura doté de
particularités physiques qui s'avéreront
spécialement adaptées à sa survie :
peut-être un cou plus long, ou peut-être des pattes
arrière plus longues, ou peut-être des excroissances sur
le front qui seront les amorces de cornes, ou peut-être . . . une
déformation du squelette qui rendra la marche bipède plus
commode. Si les conditions de l'environnement se poursuivent assez
longtemps, elles peuvent alors influencer de la même
manière un nombre suffisant d'embryons et sur un nombre
suffisant de générations pour que, finalement, une
mutation s'opère et que l'on puisse dire qu'une nouvelle
espèce est apparue.
Si l'on pense, par exemple, à l'évolution qui a
donné naissance aux premières espèces humaines, on
peut se souvenir que la vitesse du développement des embryons
est différente chez les espèces des grands singes et chez
l'humain. Peut-être est-ce là la cause initiale qui a
mené à notre évolution : simplement un
ralentissement de la vitesse du développement embryonnaire,
ralentissement qui a changé la force et donc la dynamique de sa
croissance, et par conséquent la forme de cette dynamique dans
l'espace, déplaçant d'abord juste un peu
l'équilibre du squelette pour permettre une marche debout plus
confortable et pour permettre, ensuite, le développement
possible du cerveau désormais bien en équilibre au sommet
de la colonne vertébrale.
Il y a quelques années, Anne Dambricourt-Malassé avait
suggéré que, dans l'embryon, la dynamique qui
génère l'angle entre la base du crâne et la face
évolue ainsi de générations en
générations pour, à l'arrivée, parvenir
à l'angle qui permet commodément la tenue du cou sur un
corps vertical (voir son article "Nouveau regard sur l'Origine de
l'homme" dans le numéro n° 286 d'avril 1996 de la revue La
Recherche).
Elle a été alors très injustement critiquée
pour avoir repéré que cette évolution se faisait
dans un sens précis, car on la soupçonnait de donner un
sens religieux à ce sens, comme si le fait que les choses
aillent dans un sens impliquait nécessairement qu'elles y
allaient pour atteindre un but précis et que, de plus, ce but
avait été nécessairement prévu ou voulu
d'avance.
La pierre qui tombe va dans un sens, comme la rivière qui coule,
et cela ne va pourtant vers aucun but. Dire que tout n'arrive pas par
hasard mais suit une logique propre qui se développe
progressivement, n'implique pas nécessairement que quelqu'un ou
quelqu'être supérieur a prévu cette logique, c'est
seulement dire que les choses se poussent l'une l'autre et vont ainsi
de l'avant de manière irréversible et qui se joue des
obstacles qui l'entravent.
Fondamentalement, je crois que, au-delà du cas du passage vers
l'humain qui était le but de son article, Anne
Dambricourt-Malassé a traité du point fondamental pour ce
qui concerne, de manière générale,
l'évolution des espèces : la plasticité de la
dynamique de croissance des embryons, qui fait que toutes les
transformations sont alors possibles et même, c'est le point
crucial, pratiquement inévitables, tellement, à ce stade
du développement de l'être vivant complexe, le moindre
changement peut avoir des conséquences considérables
parce qu'il sera amplifié par la croissance future, croissance
qu'il va localement contrarier ou au contraire faciliter au
détriment de la croissance du reste de l'embryon.
Ne faut-il donc pas admettre que la croissance de l'embryon est le
vivier de la transformation des espèces, que c'est là que
le hasard façonne des quantités d'essais qui auront un
sort plus ou moins réussi ? Ce ne peut être un hasard si
c'est précisément une femme (porteuse d'embryon, ne serait-ce que potentiellement) qui a, la première a
ma connaissance, proposé cette idée.
Note 1 : si vous avez jamais fait un
peu de programmation, vous devez savoir que jamais une erreur de
programmation ne vous donne un programme qui fonctionne correctement
"par hasard". Ou bien cela ne fonctionne pas du tout, ou bien cela
fonctionne mal, mais, si du moins j'en crois mon expérience
personnelle, jamais cela ne donne un programme nouveau qui, par hasard,
fait des choses surprenantes auxquelles vous n'aviez pas pensé
au départ. Je serais d'ailleurs curieux de savoir, depuis que la
programmation informatique existe, combien de fois des erreurs de
programmation ont, par hasard, produit un nouveau programme plus
fonctionnel que prévu par son programmateur, ou même tout
simplement correctement fonctionnel (retour au texte ).
Le 28 janvier 2005 (corrigé ponctuellement le 13 mai 2007)
|
accueil |
Science |
haut |
autres textes 2005 |
 |